Основные функции белков в клетке: назовите функции белков в клетке
Содержание
назовите функции белков в клетке
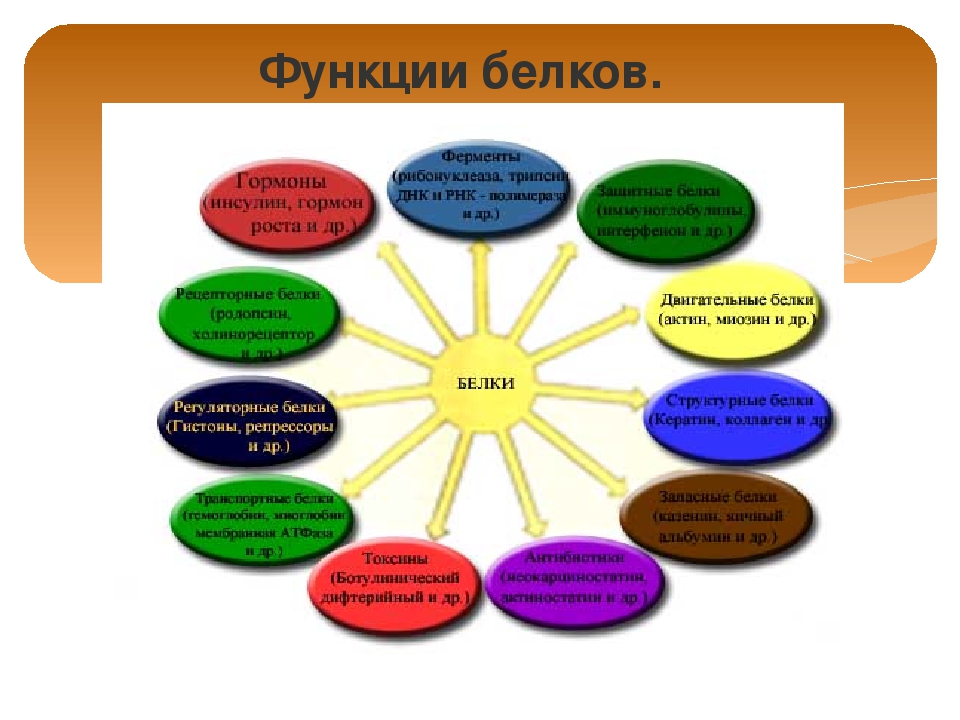
Роль белков в клетке и организме Белки играют исключительно большую роль в процессах жизнедеятельности клетки и организма, им свойственны следующие функции. Структурная. Входят в состав внутриклеточных структур‚ тканей и органов. Например, коллаген и эластин служат компонентами соединительной ткани: костей‚ сухожилий‚ хрящей; фиброин входит в состав шелка‚ паутины; кератин входит в состав эпидермиса и его производных (волосы‚ рога‚ перья) . Образуют оболочки (капсиды) вирусов. Ферментативная. Все химические реакции в клетке протекают при участии биологических катализаторов — ферментов (оксидоредуктазы, гидролазы, лигазы, трансферазы, изомеразы, и лиазы) . Регуляторная. Например, гормоны инсулин и глюкагон регулируют обмен глюкозы. Белки–гистоны участвуют в пространственной организации хроматина, и тем самым влияют на экспрессию генов. Транспортная. Гемоглобин переносит кислород в крови позвоночных, гемоцианин в гемолимфе некоторых беспозвоночных, миоглобин — в мышцах.
Сывороточный альбумин служит для транспорта жирных кислот‚ липидов и т. п. Мембранные транспортные белки обеспечивают активный транспорт веществ через клеточные мембраны (Na+, К+-АТФаза) . Цитохромы осуществляют перенос электронов по электронтранспортным цепям митохондрий и хлоропластов. Защитная. Например, антитела (иммуноглобулины) образуют комплексы с антигенами бактерий и с инородными белками. Интерфероны блокируют синтез вирусного белка в инфицированной клетке. Фибриноген и тромбин участвуют в процессах свертывания крови. Сократительная (двигательная) . Белки актин и миозин обеспечивают процессы мышечного сокращения и сокращения элементов цитоскелета. Сигнальная (рецепторная) . Белки клеточных мембран входят в состав рецепторов и поверхностных антигенов. Запасающие белки. Казеин молока, альбумин куриного яйца, ферритин (запасает железо в селезенке) .
Белки-токсины. Дифтерийный токсин. Энергетическая функция. При распаде 1 г белка до конечных продуктов обмена (СО2, Н2О, Nh4, Н2S, SО2) выделяется 17‚6 кДж или 4‚2 ккал энергии
Какова функция белков в клетке?
Белки играют исключительно большую роль в процессах жизнедеятельности клетки и организма, им свойственны следующие функции. Структурная. Входят в состав внутриклеточных структур‚ тканей и органов. Например, коллаген и эластин служат компонентами соединительной ткани: костей‚ сухожилий‚ хрящей; фиброин входит в состав шелка‚ паутины; кератин входит в состав эпидермиса и его производных (волосы‚ рога‚ перья) . Образуют оболочки (капсиды) вирусов. Ферментативная. Все химические реакции в клетке протекают при участии биологических катализаторов — ферментов (оксидоредуктазы, гидролазы, лигазы, трансферазы, изомеразы, и лиазы) . Регуляторная. Например, гормоны инсулин и глюкагон регулируют обмен глюкозы. Белки–гистоны участвуют в пространственной организации хроматина, и тем самым влияют на экспрессию генов.
Транспортная. Гемоглобин переносит кислород в крови позвоночных, гемоцианин в гемолимфе некоторых беспозвоночных, миоглобин — в мышцах. Сывороточный альбумин служит для транспорта жирных кислот‚ липидов и т. п. Мембранные транспортные белки обеспечивают активный транспорт веществ через клеточные мембраны (Na+, К+-АТФаза) . Цитохромы осуществляют перенос электронов по электронтранспортным цепям митохондрий и хлоропластов. Защитная. Например, антитела (иммуноглобулины) образуют комплексы с антигенами бактерий и с инородными белками. Интерфероны блокируют синтез вирусного белка в инфицированной клетке. Фибриноген и тромбин участвуют в процессах свертывания крови. Сократительная (двигательная) . Белки актин и миозин обеспечивают процессы мышечного сокращения и сокращения элементов цитоскелета.
Сигнальная (рецепторная) . Белки клеточных мембран входят в состав рецепторов и поверхностных антигенов. Запасающие белки. Казеин молока, альбумин куриного яйца, ферритин (запасает железо в селезенке) . Белки-токсины. Дифтерийный токсин Энергетическая функция. При распаде 1 г белка до конечных продуктов обмена (СО2, Н2О, Nh4, Н2S, SО2) выделяется 17‚6 кДж или 4‚2 ккал энергии
Функции белков в организме | Химия онлайн
Функции белков в природе универсальны. Белки входят в состав всех живых организмов. Мышцы, кости, покровные ткани, внутренние органы, хрящи, шерсть, кровь — все это белковые вещества.
Растения синтезируют белки из углекислого газа и воды за счет фотосинтеза. Животные организмы получают, в основном, готовые аминокислоты с пищей и на их базе строят белки своего организма.
Ни один из известных нам живых организмов не обходится без белков. Белки служат питательными веществами, они регулируют обмен веществ, исполняя роль ферментов – катализаторов обмена веществ, способствуют переносу кислорода по всему организму и его поглощению, играют важную роль в функционировании нервной системы, являются механической основой мышечного сокращения, участвуют в передаче генетической информации и т.д.
Видеофильм«Функции белков»
Разнообразные функции белков определяются a-аминокислотным составом и строением их высокоорганизованных макромолекул.
1. Каталитическая (ферментативная) функция
Каталитическая функция — одна из основных функций белков. Абсолютно все биохимические процессы в организме протекают в присутствии катализаторов –
ферментов. Все известные ферменты представляют собой белковые молекулы.
Белки – это очень мощные катализаторы. Они ускоряют реакции в миллионы раз, причем для каждой реакции существует свой фермент.
В настоящее время известно свыше 2000 различных ферментов, которые являются биологическими катализаторами.
Например, фермент пепсин расщепляет белки в процессе пищеварения.
Даже такая простая реакция как гидратация углекислого газа катализируется ферментом карбоангидразой.
Ферменты катализируют реакции расщепления сложных молекул (катаболизм) и их синтеза (анаболизм), а также репликации ДНК и матричного синтеза РНК.
2. Транспортная функция
Некоторые белки способны присоединять и переносить (транспортировать) различные вещества по крови от одного органа к другому и в пределах клетки.
Белки транспортируют липиды (липопротеиды), углеводы (гликопротеиды), ионы металлов (глобулины), кислород и углекислый газ (гемоглобин), некоторые витамины, гормоны и др.
Например, альбумины крови транспортируют липиды и высшие жирные кислоты (ВЖК), лекарственные вещества, билирубин.
Белок эритроцитов крови гемоглобин соединяется в легких с кислородом, превращаясь в оксигемоглобин. Достигая с током крови органов и тканей, оксигемоглобин расщепляется и отдает кислород, необходимый для обеспечения окислительных процессов в тканях.
Белок миоглобин запасает кислород в мышцах.
Специфические белки-переносчики обеспечивают проникновение минеральных веществ и витаминов через мембраны клеток и субклеточных структур.
3. Защитная функция
Защитную функцию выполняют специфические белки (антитела — иммуноглобулины), которые вырабатываются иммунной системой организма. Они обеспечивают физическую, химическую и иммунную защиту организма путем связывания и обезвреживания веществ, поступающих в организм или появляющихся в результате жизнедеятельности бактерий и вирусов.
Например, белок плазмы крови фибриноген участвует в свертывании крови (образовывает сгусток). Это защищает организм от потери крови при ранениях.
Альбумины обезвреживают ядовитые вещества (ВЖК и билирубин) в крови.
Многие живые существа для обеспечения защиты выделяют белки, называемые токсинами, которые в большинстве случаев являются сильными ядами. В свою очередь, некоторые организмы способны вырабатывать антитоксины, которые подавляют действие этих ядов.
4. Сократительная (двигательная) функция
Важным признаком жизни является подвижность, в основе которой лежит данная функция белков, таких как актин и миозин – белки мышц. Кроме мышечных сокращений к этой функции относят изменение форм клеток и субклеточных частиц.
B результате взаимодействия белков происходит передвижение в пространстве, сокращение и расслабление сердца, движение других внутренних органов.
5. Структурная функция
Структурная функция — одна из важнейших функций белков. Белки играют большую роль в формировании всех клеточных структур.
Белки – это строительный материал клеток. Из них построены опорные, мышечные, покровные ткани.
Некоторые из них (коллаген соединительной ткани, кератин волос, ногтей, эластин стенок кровеносных сосудов, фиброин шелка и др.) выполняют почти исключительно структурную функцию.
Кератин синтезируется кожей. Волосы и ногти – это производные кожи.
В комплексе с липидами белки участвуют в построении мембран клеток и внутриклеточных образований.
6. Гормональная (регуляторная) функция
Регуляторная функция присуща белкам-гормонам (регуляторам). Они регулируют различные физиологические процессы.
Например, наиболее известным гормоном является инсулин, регулирующий содержание глюкозы в крови. При недостатке инсулина в организме возникает заболевание, известное как сахарный диабет.
Интересно знать!
В плазме некоторых антарктических рыб содержатся белки со свойствами антифриза, предохраняющие рыб от замерзания, а у ряда насекомых в местах прикрепления крыльев находится белок резилин, обладающий почти идеальной эластичностью. В одном из африканских растений синтезируется белок монеллин с очень сладким вкусом.
7. Питательная (запасная) функция
Питательная функция осуществляется резервными белками, которые запасаются в качестве источника энергии и вещества.
Например: казеин, яичный альбумин, белки яйца обеспечивают рост и развитие плода, а белки молока служат источником питания для новорожденного.
8. Рецепторная (сигнальная) функция
Некоторые белки (белки-рецепторы), встроенные в клеточную мембрану, способны изменять свою структуру под воздействием внешней среды. Так происходит прием сигналов извне и передача информации в клетку.
Например, действие света на сетчатку глаза воспринимается фоторецептором родопсином.
Рецепторы, активизируемые низкомолекулярными веществами типа ацетилхолина, передают нервные импульсы в местах соединения нервных клеток.
9. Энергетическая функция
Белки могут выполнять энергетическую функцию, являясь одним из источников энергии в клетке (после их гидролиза). Обычно белки расходуются на энергетические нужды в крайних случаях, когда исчерпаны запасы углеводов и жиров.
При полном расщеплении 1 г белка до конечных продуктов выделяется 17,6 кДж энергии. Но в качестве источника энергии белки используются крайне редко. Аминокислоты, высвобождающиеся при расщеплении белковых молекул, используются для построения новых белков.
Белки
Белки
Белок – это фундамент организма. Он целиком построен из маленьких строительных блоков, называемых аминокислотами. Аминокислоты являются основными составными частями и строительными элементами белковой молекулы.
Название «протеины» (от греч. protos – первый, важнейший) более точно отражает первостепенное биологическое значение этого класса веществ, хотя в отечественной литературе их принято называть белками или белковыми веществами по аналогии с белком куриного яйца, приобретающего при кипячении (денатурации) белый цвет.
Молекула белка построена из 100 или более остатков аминокислот, ковалентно связанных в полимерные цепи. В человеческом организме 5 миллионов белков, причем ни один из белков человека не идентичен с белком любого другого живого организма.
Несмотря на такое разнообразие белковых структур для их построения необходимы всего 22 аминокислоты, 9 из которых незаменимы, то есть должны поступать с пищей человека, они не синтезируются в организме человека, остальные аминокислоты могут образовываться в нашем организме из других аминокислот. Таким образом, необходимо обеспечить адекватную поставку организму этих аминокислот соответствующим питанием с хорошо сбалансированным составом животных и растительных белков.
Большинство растительных белков, даже очень важных, содержит лишь незначительное число незаменимых аминокислот – именно поэтому строгие вегетарианцы (не потребляющие даже яиц и молочных продуктов) могут испытывать дефицит данных аминокислот. Невозможно переоценить важность сбалансированного питания для обеспечения достаточного уровня всех незаменимых аминокислот. Это существенный фактор здоровья!
Биологические функции белков очень разнообразны. Они выполняют каталитические, регуляторные, структурные, двигательные, транспортные, защитные, запасные и другие функции. Белки составляют основу биомембран, важнейшей составной части клетки и клеточных компонентов.
Исключительное свойство белков – самоорганизация структуры, то есть способность самопроизвольно создавать определенную свойственную только данному белку структуру. Для того, чтобы организм мог эффективно использовать и синтезировать белок, должны присутствовать все незаменимые аминокислоты и в необходимой пропорции. Даже временное отсутствие одной незаменимой аминокислоты может отрицательно сказаться на синтезе белка. При уменьшении количества любой незаменимой аминокислоты или ее отсутствии пропорционально уменьшается эффективность всех остальных аминокислот.
Девять незаменимых аминокислот – гистидин (необходим для детей), изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан и валин. Организм в состоянии строить белки только при наличии достаточных количеств их всех. При отсутствии хотя бы одной из этих кислот белки уже не синтезируются, а пища используется как источник энергии или сохраняется в жировых отложениях.
Остальные 13 аминокислот синтезируются в организме человека в реакциях трансаминации. Эти аминокислоты – аланин, аргинин, аспарагин, карнитин, цистеин, цистин, глутаминовая кислота, глутамин, глицин, гидроксипролин, пролин, серин, тирозин.
Помимо 22 аминокислот, входящих в состав белков, известно свыше 150 других, которые встречаются в различных клетках и тканях либо в свободном, либо в связанном виде, но никогда не встречаются в составе белков.
Значение аминокислот не ограничивается их ролью в синтезе тканевых белков. Каждая из них выполняет в организме свои особые функции, направленные на поддержание гомеостаза организма.
Лишение всех аминокислот приводит к совершенной потере белка организмом, истощению, полной потере энергии, потере веса, анемии, разрушению мышц, в самом тяжелом случае – к смерти.
ФУНКЦИИ БЕЛКОВ
Белки выполняют множество самых разнообразных функций, характерных для живых организмов. Здесь будут перечислены главные и в некотором смысле уникальные для жизнедеятельности человека функции, не свойственные или лишь частично присущие другим классам биополимеров.
Каталитическая функция белков. Все до сих пор открытые биологические катализаторы – ферменты являются белками. Эта функция белков является уникальной, не свойственной другим полимерным молекулам.
Питательная (резервная) функция белков. К таким белкам относятся так называемые резервные белки, являющиеся источниками питания для развития плода; белки яйца (овальбумины) и основной белок молока (казеин) также выполняют главным образом питательную функцию. Ряд других белков несомненно используется в организме в качестве источника аминокислот, которые в свою очередь являются предшественниками биологически активных веществ, регулирующих процессы обмена веществ.
Транспортная функция белков. Дыхательная функция крови, в частности перенос кислорода, целиком осуществляется молекулами гемоглобина – белка эритроцитов. В транспорте липидов принимают участие альбумины сыворотки крови. Ряд других сывороточных белков образует комплексы с жирами, медью, железом, тироксином, витамином A и другими соединениями, обеспечивая их доставку в соответствующие органы-мишени.
Защитная функция белков. Основную функцию защиты в организме выполняет иммунологическая система, которая обеспечивает синтез специфических защитных белков – антител в отвег на поступление в организм бактерий, токсинов или вирусов. Высокая специфичность взаимодействия антител с антигенами (чужеродными веществами) по типу белок – белковое взаимодействие способствует нейтрализации их биологического действия и сохранению нормального состояния. В качестве другого примера защитной роли можно привести способность ряда белков крови к свертыванию. Свертывание белка плазмы крови фибриногена приводит к образованию сгустка крови, что предохраняет от потери крови при ранениях.
Сократительная функция белков. В акте мышечного сокращения и расслабления участвует множество белковых веществ тела. Однако главную роль в этих жизненно важных процессах играют актин и миозин – специфические белки мышечной ткани. Сократительная функция присуща не только мышечным белкам, но и белкам ряда субклеточных структур, что обеспечивает тончайшие процессы жизнедеятельности клеток.
Структурная функция белков. Данная функция белков многопланова. Белки со структурными функциями занимают по количеству первое место среди других белков тела человека. Широко распространены такие важные структурные белки, как коллаген в соединительной ткани, кератин в волосах, ногтях, коже, эластин в сосудистой стенке и др. Не менее важную роль выполняют белки в комплексе с углеводами в формировании ряда секретов – мукоидов, муцинов и т. д. Наконец, в комплексе с липидами (в частности, фосфолипидами) белки участвуют в образовании биомембран клеток.
Гормональная функция белков. Обмен веществ в организме регулируется разнообразными механизмами. В этой регуляции важное место занимают гормоны, вырабатываемые в железах внутренней секреции. Ряд гормонов представлен белками или полипептидами, например, гормоны гипофиза, поджелудочной железы и др.
Можно указать еще на некоторые другие жизненно важные функции белков, в частности, на способность белков к сохранению онкотического давления в клетках и в крови, на буферные свойства белков, регулирующие физиологическое значение pH внутренней среды, и др.
Строение и функции белков — конспект
Вернуться к теме «Строение и функции белков»
Белки – полимеры, мономерами которых являются аминокислоты.
Среди органических веществ белки занимают первое место по количеству и по значению. В организме человека встречаются 5 млн разнообразных белковых молекул, отличающихся не только друг от друга, но и от белков других организмов. Несмотря на такое разнообразие и сложность строения они построены всего из 20 различных аминокислот.
Строение аминокислоты:
В левой части молекулы расположены группа h3N–, которая обладает свойствами основания; справа — группа –COOH — кислотная, характерная для всех органических кислот. Следовательно, аминокислоты – амфотерные соединения, совмещающие свойства и кислоты и основания. Этим обусловлена их способность взаимодействовать друг с другом. Соединяясь, молекулы аминокислот образуют связи между углеродом кислотной и азотом основной групп. Такие связи называются ковалентными, а в данном случае – пептидными связями:
Соединение двух аминокислот в одну молекулу называется дипептидом, трех аминокислот – трипептидом и т. д., а соединение, состоящее из 20 и более аминокислотных остатков, – полипептидом.
Последовательность аминокислот в полипептидной цепи принято называть первичной структурой белка.
Однако молекула белка в виде цепи аминокислотных остатков, последовательно соединенных между собой пептидными связями, еще не способна выполнять специфические функции. Для этого необходима более высокая структурная организация. Путем образования водородных связей между остатками карбоксильных и аминогрупп разных аминокислот белковая молекула принимает вид спирали (α-структура) или складчатого слоя – «гармошки» (β-структура). Это вторичная структура белка. Но и ее часто недостаточно для приобретения характерной биологической активности.
Часто только молекула, обладающая третичной структурой, может выполнять роль катализатора или любую другую. Третичная структура образуется благодаря взаимодействию радикалов, в частности радикалов аминокислоты цистеина, которые содержат серу. Атомы серы двух аминокислот, находящихся на некотором расстоянии друг от друга в полипептидной цепи, соединяются, образуя так называемые дисульфидные, или S–S, связи. Благодаря этим взаимодействиям, а также другим, менее сильным связям, белковая спираль сворачивается и приобретает форму шарика, или глобулы. Способ укладки полипептидных спиралей в глобуле называют третичной структурой белка. Многие белки, обладающие третичной структурой, могут выполнять свою биологическую роль в клетке. Однако для осуществления некоторых функций организма требуется участие белков с еще более высоким уровнем организации.
Такую организацию называют четвертичной структурой. Присутствует не у всех белков. Она представляет собой функциональное объединение нескольких (двух, трех и более) молекул белка, обладающих третичной структурной организацией. Пример такого сложного белка – гемоглобин. Его молекула состоит из четырех связанных между собой молекул. Другим примером может служить гормон поджелудочной железы – инсулин, включающий два компонента. В состав четвертичной структуры некоторых белков включаются помимо белковых субъединиц и разнообразные небелковые компоненты. Тот же гемоглобин содержит сложное гетероциклическое соединение, в состав которого входит железо.
Строение белковой молекулы: А – первичная; Б – вторичная; В – третичная; Г – четвертичная структура
Строение молекулы гемоглобина
Гемоглобин – белок четвертичной структуры. В молекуле гемоглобина белковый компонент представлен белком глобином, небелковый компонент – гем. Глобин состоит из 4 субъединиц. Внутри каждой субъединицы имеется гидрофобный «карман», в котором располагается гем. Содержащийся в геме атом железа связывает кислород.
Свойства белка
Белки, как и другие неорганические и органические соединения, обладают рядом физико-химических свойств:
Белки – преимущественно водорастворимые молекулы и, следовательно, могут проявлять свою функциональную активность только в водных растворах.
Белковые молекулы несут большой поверхностный заряд. Это определяет целый ряд электрохимических эффектов, например изменение проницаемости мембран каталитической активности и других функций.
Белки термолабильны, то есть проявляют свою активность в узких температурных рамках.
Денатурация и ренатурация белков
Денатурация – это утрата белковой молекулой своей структурной организации: четвертичной, третичной, вторичной, а при более жестких условиях – и первичной структуры. В результате денатурации белок теряет способность выполнять свою функцию. Причинами денатурации могут быть высокая температура, ультрафиолетовое излучение, действие сильных кислот и щелочей, тяжелых металлов и органических растворителей. Если изменение условий среды не приводит к разрушению первичной структуры молекулы, то при восстановлении нормальных условий среды полностью воссоздается структура белка и его функциональная активность. Такой процесс носит название ренатурации.
Функции белков
1. Каталитическая (ферментативная) функция:
Многие белки являются ферментами. Ферменты — это биологические катализаторы, т. е. вещества, ускоряющие протекание химических реакций в живых организмах. Ферменты участвуют в процессах синтеза и расщепления различных веществ. Они обеспечивают фиксацию углерода в процессе фотосинтеза, расщепление питательных веществ в пищеварительном тракте и т. д.
2. Транспортная функция
Многие белки способны присоединять и переносить различные вещества. Гемоглобин связывает и переносит кислород и углекислый газ. Альбумины крови транспортируют жирные кислоты, глобулины — ионы металлов и гормоны. Многие белки, входящие в состав цитоплазматической мембраны, участвуют в транспорте веществ в клетку и из нее.
3. Защитная функция
Белки предохраняют организм от вторжения чужеродных организмов и от повреждений. Так, в ответ на проникновение чужеродных объектов (антигенов) определенные лейкоциты вырабатывают специфические белки — иммуноглобулины (антитела), участвующие в иммунном ответе организма. Белок плазмы крови фибриноген, участвуя в свертывании крови и тем самым уменьшая кровопотери.
4. Двигательная (сократительная) функция
Сократительные белки обеспечивают способность клеток, тканей, органов и целых организмов изменять форму, двигаться. Так, актин и миозин обеспечивают работу мышц и немышечные внутриклеточные сокращения.
5. Структурная (строительная, пластическая) функция
Белки входят в состав всех клеток и тканей живых организмов. Белки являются обязательным компонентом всех клеточных мембран и органоидов клетки. Из белков построены элементы цитоскелета, сократительные элементы мышечных волокон. Преимущественно из белков состоят хрящи и сухожилия. В их состав входит белок коллаген. Важнейшим структурным компонентом перьев, волос, ногтей, когтей, рогов, копыт у животных является белок кератин. В состав связок, стенок артерий и лёгких входит структурный белок эластин.
6. Сигнальная (рецепторная) функция
Некоторые белки клеточных мембран способны изменять свою структуру в ответ на действие внешних факторов. С помощью этих белков происходит прием сигналов из внешней среды и передача информации в клетку.
7. Регуляторная функция
Некоторые белки являются гормонами. Они влияют на различные физиологические процессы. Например, инсулин и глюкагон регулируют содержание глюкозы в крови, а соматотропин (гормон роста) — процессы роста и физического развития.
8. Запасающая (питательная) функция
В семенах растений запасаются резервные белки, которые используются при прорастании зародышем.
9. Энергетическая функция
При полном окислении 1 г белка выделяется 17,6 кДж энергии. Однако белки расходуются на энергетические нужды лишь в крайних случаях, когда исчерпаны запасы углеводов и жиров.
Найдены неожиданные функции белка, ответственного за программируемую гибель клеток
Семейство белков-каспаз участвует как в запуске, так и в усилении процесса апоптоза в ответ на различные повреждения клетки. Белок каспаза-2 – его самый эволюционно консервативный представитель, возникший довольно давно и мало изменившийся с тех пор. Но в последние годы стали появляться данные о том, что каспаза-2 участвует во множестве других процессов, не связанных напрямую с апоптозом. Так, этот белок может подавлять развитие раковых опухолей или регулировать обмен веществ в клетке.
«Для анализа потенциально возможной связи каспазы-2 с другими белками мы использовали хорошо известную дрожжевую двугибридную систему. С помощью этого метода мы получили достаточно большой список белков, которые могут связываться с каспазой-2. Аспирант лаборатории Алексей Замараев проверил этот список путем биоинформатического анализа и ранжировал их по вероятности взаимодействия. Благодаря такому анализу после детального биохимического исследования мы некоторое время назад нашли новый взаимодействующий с каспазой-2 регуляторный фактор», – рассказал о работе руководитель научной группы и ведущий автор статьи Борис Животовский, доктор биологических наук и руководитель лаборатории исследования механизмов апоптоза факультета фундаментальной медицины МГУ имени М.В. Ломоносова.
Этот регуляторный фактор, найденный во время одного из предыдущих исследований, называется RFXANK. Он участвует в управлении работой гена, контролирующего молекулы главного комплекса гистосовместимости, которые помогают специальным клеткам иммунной системы различать «своих» и «чужаков», защищая организм от инфекций.
Продолжив поиски, ученые обнаружили еще один белок, с которым взаимодействует каспаза-2. Обозначают его английской аббревиатурой FAN, а его полное название – активатор нейтральной сфингомиелиназы. Он управляет производством церамидов – важных компонентов многих липидов (в просторечии часто называемых жирами, хотя жиры – более узкая категория, нежели липиды). Кроме того, FAN контролирует миграцию клеток и производство интерлейкина-6 – молекулы, которая регулирует воспалительные процессы. Другая важная функция FAN – руководить образованием везикул, маленьких внутриклеточных пузырьков, состоящих из того же белково-липидного двухслойного «материала», что и клеточная мембрана. В таких пузырьках различные молекулы могут перемещаться по клетке, «перевариваться» с помощью специальных белков или выбрасываться наружу.
Определить, на какие из этого множества ролей белка FAN влияет каспаза-2, помог современный биологический метод – липидомика. В отличие от своих более популярных «сестер»: протеомики, которая занимается взаимодействиями белков (протеинов), и геномики, изучающей гены и геномы, — липидомика помогает построить сети реакций, в которые вовлечены липиды. Именно эти реакции были важны для понимания работы FAN, функции которого так тесно с ними связаны.
«Совместная работа исследователей МГУ и Каролинского института (Швеция) позволила, используя липидомику, исключить участие каспазы-2 в образовании церамидов, но установить, что взаимодействие каспазы-2 с FAN отвечает за регулирование выхода из клетки интерлейкина-6, размер везикул и скорость миграции клеток, – суммирует выводы исследования Борис Животовский. – Последнее принципиально важно для понимания процесса метастазирования опухоли».
На первый взгляд кажется непонятным, какое отношение FAN может иметь к образованию метастаз – вторичных очагов роста опухоли. Но миграция клеток, которую упоминает ученый, такой же двоякий процесс, как и усиленное их деление, пролиферация. С одной стороны, оба они важны для роста тканей и заживления ран. Но ускоренные без надобности пролиферация и миграция могут сослужить организму и плохую службу, помогая раковой опухоли расти и добираться в новые и новые части организма. В ходе исследования ученые экспериментально доказали, что каспаза-2 через взаимодействие с FAN не ускоряет деление клеток, но помогает им двигаться быстрее. Теоретически, если научиться правильно выключать взаимодействие каспазы-2 и FAN, то можно замедлить процесс образования метастаз, а его включение помогло бы улучшить заживление ран.
Перед применением этого подхода на практике нужно удостовериться, что другие многочисленные функции обоих белков не слишком пострадают от подобного лечения. Как мы помним, открытое в ходе этой же работы взаимодействие каспазы-2 с FAN влияет и на формирование везикул. Подавив действие каспазы-2, ученые убедились, что без этого белка внутриклеточные пузырьки сильно увеличивались в размере. Такая особенность везикул уже была ранее известна как проявление синдрома Чедиака-Хигаши, при котором у человека возникает светобоязнь, нервный тик и нарушения работы иммунной системы. Поэтому только дальнейшие исследования покажут, можно ли бороться с метастазами через подавление связывания каспазы-2 с FAN без вреда для других функций обоих белков.
Свойства функции белков
При изучении темы поставлены задачи о
формировании знаний о важнейших свойствах и
функциях белков. Проводится практическая работа
по наблюдению расщепления пероксида водорода
ферментом каталазой. Повторяется материал, и
контролируются знания учащихся.
Урок проводится в два этапа. На первом
этапе в ходе беседы и рассказа учитель объясняет
функции белков, демонстрируя таблицы по общей
биологии. Объясняется значение каталазы в
растительной и животной клетке, ее функции –
расщепление опасно для клетки Н2О2 до
кислорода и воды.
На втором этапе проводится
практическая работа по обнаружению
каталитической функции каталазы и явления
денатурации. Учащимися делается общий вывод
после окончания работы.
В конце урока подводятся итоги,
выставляются отметки, закрепляется новая тема,
выясняются трудные вопросы и задается домашнее
задание.
Тема: Свойства и функции белков.
Задачи:
Сформулировать знания о важнейших свойствах и
функциях белков.
Провести практическую работу по наблюдению
расщепления пероксида водорода ферментом
каталазой.
Повторить материал и проконтролировать знания
учащихся по теме “Строение белков”.
Оборудование:
1. Раздаточный материал:
1) Штатив с тремя пробирками. В первой
пробирке кусочек сырого мяса, во второй пробирке
кусочек сырого картофеля, в третьей пробирке
кусочек вареного картофеля.
2) Склянка с 3% раствором Н2О2.
3) Баночки с элодеей.
4) Микроскопы и покровные стекла.
2. Демонстрационный материал:
1) Таблицы по общей биологии.
2) Кодограмма.
3) Тесты.
Ход урока
I. Повторение и опрос домашнего задания
проводятся несколькими методами:
1) Работа у доски (Приложение №2).
2) Компьютерное тестирование (Приложение №3).
3) Устное повторение по вопросам:
– Что такое белок?
– Что такое липиды?
– Какие вещества образуют основу клеточной
мембраны?
– Какую функцию выполняют углеводы?
– Из чего состоят белки?
II. Изучение нового материала.
Учитель использует таблицы по общей биологии.
На первом этапе в ходе беседы учитель объясняет
все функции белков. Объясняется значение
каталазы в растительной и животной клетке: ее
присутствие во всех клетках растений и животных,
ее функции – расщепление опасного для клетки
пероксида водорода до кислорода и воды.
Рассказ учителя:
Аминокислотный состав, структура белковой
молекулы определяют его свойства. Белки являются
амфотерными соединениями, сочетают в себе
основные и кислотные свойства. Чем больше
кислотных аминокислот в белке, тем ярче выражены
его кислотные свойства.
Есть белки растворимые, есть нерастворимые,
выполняющие механические функции (фиброин,
кератин, коллаген). Есть белки необычайно
активные (ферменты), есть химически неактивные,
устойчивые к воздействию различных условий
среды и крайне неустойчивые.
Внешние факторы могут вызывать структурные
организующие молекул белки. Процесс утраты
трехмерной конформации, присущей данной
молекуле белка, называют денатурацией.
Учитель демонстрирует таблицы, где показаны
структуры белка.
Денатурация может быть: обратимой и
необратимой, называются причины.
Благодаря сложности разнообразию форм и
состава белки играют важную роль в
жизнедеятельности клетки и организма в целом,
функции их разнообразны:
Строительная.
Белки участвуют в образовании
клеточных и внеклеточных структур, входят в
состав клеточных мембран шерсти, волос,
сухожилий, стенок сосудов и т.д.
Транспортная.
Некоторые белки способны
присоединять различные вещества и переносить их
к различным тканям и органам тела(гемоглобин).
Регуляторная.
Большая группа белков
организма принимает участие в регуляции
процессов обмена веществ. Такими белками
являются гормоны – биологически активные
вещества. Гормон инсулин регулирует уровень
сахара в крови путем повышения проницаемости
клеточных мембран для глюкозы, способствует
синтезу гликогена, увеличивает образование
жиров из углеводов.
Защитная.
В ответ на проникновение в организм
чужеродных белков или микроорганизмов
образуются особые белки – антитела, способные
связывать и обезвреживать их. Фибрин,
образующийся из фибриногена, способствует
остановке кровотечений.
Двигательная.
Особые сократительные белки
участвуют во всех видах движений клетки и
организма.
Запасающая.
Благодаря белкам, в организме
могут откладываться про запас некоторые
вещества. К запасным белкам относятся белки яйца,
белки мозга, белки сои.
Энергетическая.
Белки являются одним из
источников энергии в клетке, они используются
тогда, когда другие источники израсходованы.
Каталитическая.
Одна из важнейших функций
белков. В состав клеток входит большое
количество веществ химически мало активных. Тем
не менее, все биохимические реакции протекают с
огромной скоростью, благодаря участию в них
биокатализаторов – ферментов, веществ белковой
природы. На скорость реакции влияет также
концентрация субстрата и концентрация фермента.
III. Практическая работа по обнаружению
каталитической функции каталазы и явления
денатурации.
Задания к практической работе:
Прилейте по 2 мл пероксида водорода в пробирки с
кусочком сырого мяса, с сырым картофелем, вареным
картофелем. Объясните наблюдаемые вами явления
при действии Н2О2 на живые и мертвые
клетки (вареный картофель).
На предметное стекло в каплю воды положите лист
элодеи и рассмотрите под микроскопом на малом
увеличении место отрыва листа от стебля.
Нанесите на лист элодеи две капли пероксида
водорода, накройте покрывным стеклом и под
микроскопом рассмотрите место отрыва листа от
стебля. Объясните бурное выделение пузырьков О2
из поврежденных клеток элодеи.
Сделать общий вывод из проделанной
практической работы и записать в тетрадь.
Итоги урока: Учитель подводит итоги урока,
выставляет отметки учащимся, выясняет, что
узнали нового, что было трудно в новой теме, как
найти ответы на вопросы – параграф 27.
Домашнее задание: – параграф 27,
– закончить вывод по практической работе.
Приложение
Многофункциональность белка: принципы и механизмы
Структурная сложность и компактность белка могут быть достигнуты с помощью различных механизмов, действующих на уровне генома, транскриптома и протеома (). Несколько примеров многофункциональных белков, описанных ниже, могут продемонстрировать, как эти механизмы могут развить многофункциональность.
Уровень генома
Основными инструментами создания биологического разнообразия на уровне генома являются мутации (вставка или удаление нуклеотидов в последовательности ДНК), транспозиция частей ДНК и дупликация генов с последующими генетическими расхождениями в каждом дублированном гене.Если эти изменения в генетической информации вызывают изменения в структуре и функции белков, они подвергаются эволюционному отбору и могут быть прослежены филогенетически ().
1. Мутации как источники многофункциональности
Продуктом мутировавшего гена может быть «либо альтернативный белок с эквивалентной функцией — нейтральная мутация », либо белок, который выполняет ту же функцию, но с измененной скоростью или специфичностью. профиль. Это может быть белок с измененной функцией или белок, который не функционирует — или даже сворачивается вообще »(Lesk, 2001).Например, гемоглобины взрослого человека и плода различаются заменой His на Ser (143β). В первую очередь, в результате этой мутации фетальный гемоглобин демонстрирует более низкое сродство, чем взрослый гомолог, к регуляторному лиганду дифосфоглицерату. Это способствует передаче кислорода плоду через плаценту (Lesk, 2001).
Другой пример демонстрирует роль незначительных генетических изменений в индукции фундаментальных функциональных изменений в гомологичных белках. Гаптоглобин — гомолог химотрипсина.В результате мутаций гена homologuos он потерял протеолитическую активность, характерную для химотрипсина, но получил несколько других функций: гаптоглобин может действовать как шаперон, а также как фактор, предотвращающий потерю железа из эритроцитов и как опосредующий белок иммунные ответы (Lesk, 2001).
2. Дупликация генов как источник генетических новинок
Было высказано предположение, что дупликация генов является одним из важных шагов к возникновению генетических новинок (Gu et al. 2002; Ohno, 1970).Следующие примеры могут подтвердить это предположение. GroEL — это белок теплового шока 60 кДа, широко распространенный в бактериях. Интересно, что GroEL в Chlamydiae стал дублированным в происхождении линии Chlamydiae , представляющей три различных молекулярных шаперона, а именно исходный белок GroEL1 и его паралогичные белки GroEL2 и GroEL3 (Karunakaran et al. 2003; McNally and Fares, 2007 ). Виды копий белка GroEL функционально разошлись после событий дупликации генов.Функциональная дивергенция произошла в важных функциональных областях этих белков. Несмотря на то, что три белка GroEL Chlamidiae демонстрируют значительную консервацию аминокислотной последовательности в важных областях, участвующих в связывании полипептидов, по сравнению с GroEL из Escherichia coli , значительные различия были обнаружены в областях связывания GroEL и в областях, участвующих в связывании АТФ и гидролиз. Большинство аминокислотных замен, которые повлияли на взаимодействие с белками-партнерами и были ответственны за функциональную дивергенцию между паралогами GroEL, были исправлены адаптивной эволюцией после событий дупликации гена groEL (McNally and Fares, 2007).
Стромальные взаимодействующие молекулы (STIM) представляют собой другой пример, демонстрирующий роль дупликации генов в функциональной дивергенции гомологичных белков. STIM функционируют как датчик Ca 2+ для обнаружения изменений содержания Ca 2+ во внутриклеточных хранилищах. STIM представляют собой однопролетные мембранные белки с неспаренным N-концевым EF-связывающим доменом Ca 2+ , критичным для сенсорной функции Ca 2+ . Кроме того, STIM содержат домен N-концевого стерильного α-мотива (SAM) и C-концевой (цитоплазматический) домен ERM со спиральной спиралью (Liou et al.2005; Roos et al. 2005; Williams et al. 2001; Zhang et al. 2005). STIM человека и STIM беспозвоночных имеют несколько общих функционально важных белковых доменов, но существенно различаются по С-концу.
Филогения и анализ последовательностей выявили раннюю адаптацию С-концевого дивергентного домена у Urochordata до экспансии STIMs у позвоночных. Впоследствии STIM были подвергнуты одному раунду дупликации генов уже в линии Euteleostomi у позвоночных, со вторым раундом дупликации специфичных для рыб генов. После дупликации молекулы STIM-1 и STIM-2, по-видимому, подверглись очищающей селекции. Кроме того, анализ последовательности связывающего домена Ca 2+ EF-hand и домена SAM вместе с функциональными исследованиями выявил критические области / остатки, которые, вероятно, лежат в основе функциональных изменений, и подтвердил доказательства гипотезы о том, что STIM-1 и STIM-2 могут развили отличные функциональные свойства после дупликации гена (Cai, 2007).
Уровень транскриптома
Альтернативный сплайсинг — это обычный посттранскрипционный процесс в эукариотических организмах, который производит множество изоформ транскриптов из одного гена (Black, 2003).Альтернативный сплайсинг и дупликация генов — два источника протеомного функционального разнообразия (). Согласно «независимой модели» альтернативный сплайсинг и дупликация генов являются двумя независимыми механизмами увеличения протеомной сложности. Альтернативно, «модель разделения функций» утверждает, что некоторые протеомные компоненты могут выполняться либо альтернативно сплайсированными генами, либо дублированными генами [цитируется (Su et al. 2006)].
Альтернативный сплайсинг может генерировать множественные транскрипты, кодирующие белки с тонкими или противоположными функциональными различиями, которые могут иметь серьезные биологические последствия.Недавние оценки, основанные на анализе тегов экспрессируемых последовательностей (ESTs), предполагают, что транскрипты из 35% генов человека являются альтернативным сплайсингом (Hanke et al. 1999; Mironov et al. 1999). Однако, как указал Б. Грейвли, «это число, вероятно, будет заниженным» (Graveley, 2001). Во-первых, коллекция EST человека не представляет все последовательности, кодирующие белок, и для большинства генов EST покрывают только часть транскрипта. Поскольку большая часть функционально значимого альтернативного сплайсинга происходит в кодирующей области транскрипта, многие альтернативные события сплайсинга могут быть упущены из виду при сравнении EST.Во-вторых, что, возможно, более важно, многие события альтернативного сплайсинга очень редки и происходят только в конкретной ткани в определенное время развития и / или при определенных физиологических условиях. Эти типы событий склейки, вероятно, не будут представлены в коллекциях EST (Graveley, 2001). Некоторые гены производят несколько сплайсированных мРНК, в то время как оказывается, что некоторые другие гены фактически кодируют транскрипты, которые альтернативно сплайсируются для получения десятков тысяч различных мРНК (Schmucker et al.2000).
Белки нейрексина представляют собой пример, демонстрирующий роль альтернативного сплайсинга в получении различных функций продуктами одного гена. Нейрексины представляют собой семейство нейральных белков, присутствующих у позвоночных, которые выполняют важные функции как рецепторы для нейропептидов (Missler and Sudhof, 1998) и как молекулы адгезии, которые участвуют в синаптогенезе (Scheiffele et al. 2000). Было показано, что более 1000 различных изоформ мРНК нейрексина потенциально могут быть синтезированы за счет использования альтернативных промоторов и альтернативного сплайсинга (Ullrich et al.1995). Белки, кодируемые этой альтернативной сплайсированной мРНК, обладают измененной специфичностью для своих лигандов (Ichtchenko et al. 1995; Sugita et al. 1999). Кроме того, разнообразие белков нейроксинов может определять нейронные связи (Missler et al. 1998). Было показано, что взаимодействия β-нейрексинов, присутствующих в пресинаптических клетках, с нейролигинами на поверхности постсинаптических клеток достаточно для запуска образования синапсов (Scheiffele et al. 2000). Важно отметить, что это взаимодействие происходит только в том случае, если β-neurexin кодируется мРНК, лишенной альтернативного экзона 20.Белки, синтезированные из транскриптов, содержащих экзон 20, не взаимодействуют с нейролигинами (Ichtchenko et al. 1995). Таким образом, альтернативный сплайсинг транскриптов нейрексина может играть прямую роль в контроле образования и поддержания синапсов.
Уровень протеома
Протеом — это набор белков, полученных путем трансляции мРНК, экспортируемых из ядра в цитоплазму. Трансляция мРНК в белок представляет собой заключительный этап в пути экспрессии генов, который опосредует формирование протеома из геномной информации (Gebauer and Hentze, 2004). Как мы уже подчеркивали, на уровне генома максимальный объем генетической информации должен быть закодирован в минимальном пространстве ДНК. На уровне транскриптома скрытое разнообразие будущих белков, кодируемых в ДНК и мРНК, выявляется перетасовкой различных фрагментов первичного транскрипта во время альтернативного процесса сплайсинга. Использование альтернативного промотора, в свою очередь, раскрывает разнообразие генетической информации, содержащейся в мРНК и предназначенной для трансляции. На уровне протеома максимум структурных и функциональных потенциалов, содержащихся в генетической информации, встроенной в мРНК, может быть вызван трансляционными механизмами.Последние данные, полученные путем секвенирования генома человека, показали, что геном человека состоит из примерно 35 000 генов, которые могут экспрессировать примерно 100 000 белков. Такой выброс белка возможен только потому, что существуют многочисленные механизмы, с помощью которых функционально разнообразные полипептиды продуцируются из одного гена (Touriol et al. 2003).
Способность продуцировать разные полипептиды путем трансляции одной и той же молекулы мРНК сама по себе установлена в механизме трансляции. Концептуально процесс трансляции можно разделить на три стадии — инициацию, удлинение и завершение.Во время инициации рибосома повторно собирается на мРНК, с инициаторной тРНК, связанной с сайтом Р рибосомы, и основанием, спаренным со стартовым кодоном мРНК. Элонгация относится к полимеризации пептида, основной функции рибосомы, в то время как терминация включает последовательность событий после распознавания стоп-кодона до разборки рибосомы на субъединицы и последующей диссоциации факторов, тРНК и мРНК ( Туриол и др., 2003). Механизмы, действующие на каждом этапе, определяют появление разных изоформ одного и того же генного продукта ().
A. Стадия инициации
1. Cap-зависимая инициация
Инициирование трансляции мРНК эукариот обычно происходит с помощью механизма сканирования (Kozak, 2002). Согласно модели линейного сканирования, 40S рибосомная субъединица связывается со структурой кэпа (m7GpppN) на 5′-конце мРНК и сканирует вниз по течению до первого в рамке считывания инициирующего кодона AUG (Pestova and Hellen, 1999). Канонические механизмы сканирования () управляют инициацией большинства мРНК, но также описаны три неклассических механизма инициации, зависящих от кэп: протекающее сканирование, рибосомное шунтирование и инициация терминации (Lopez-Lastra et al.2005).
a) Сканирование с утечкой
Было показано, что контекст нуклеотида вблизи стартового кодона важен для его взаимодействия с рибосомой. Если контекст не является оптимальным, некоторые рибосомы не распознают 5′-концевой проксимальный кодон AUG как начальный, пропускают его и начинают трансляцию со следующего AUG с помощью механизмов, известных как «протекающее сканирование» (Kochetov, 2006; Kozak, 2002). Этот механизм сканирования с утечкой () производит изоформы белка с альтернативными сайтами инициации аминогруппы.Следовательно, гены могут продуцировать несколько версий кодируемых белков, а в более коротких версиях, инициированных нижележащими стартовыми кодонами в рамке считывания, отсутствуют N-концевые аминокислоты полноразмерной версии изоформы (усеченная на N-конце версия) ( Туриол и др. , 2003).
Помимо дополнительных кодонов AUG, расположенных ниже основного стартового кодона, многие мРНК содержат расположенные выше AUG (uAUG), которые также могут быть выбраны трансляционными механизмами для синтеза новых изофункциональных вариантов белка.Кочетов и др. (Кочетов и др., 2005) проанализировали более 12000 мРНК человека, несущих uAUG, и обнаружили, что 3% мРНК содержат uAUG в той же рамке считывания, что и кодирующие последовательности, без промежуточных стоп-кодонов. Это означает, что такие мРНК могут кодировать дополнительные варианты белка, удлиненные на N-концах. Сравнение их предсказанного субклеточного местоположения показало, что 31% таких N-удлиненных белков имели другое местоположение, и в 19% случаев у них были сигналы секреции, отсутствующие в аннотированных вариантах (Кочетов и др.2005).
Поскольку последовательности, определяющие нацеливание на клеточные компартменты, обычно расположены в N-концевой области белка, выбор альтернативных кодонов инициации трансляции может использоваться клеткой для контроля клеточной локализации изоформ (Cai et al. 2006; Gandre и др., 2003 г .; Пакхэм и др., 1997 г.). Cai et al. (Cai et al. 2006) показали, что 85.7% событий альтернативной трансляции генерируют биологическое разнообразие, приписываемое различным субклеточным локализациям и содержанию различных доменов в альтернативных изоформах.Несколько примеров, представленных ниже, могут проиллюстрировать, как механизм сканирования с утечкой используется клеткой для увеличения разнообразия изоформ белка, продуцируемых одним геном.
Ген ДНК-лигазы 1 арабидопсиса (AtLIG1) необходим для жизнеспособности клеток. AtLIG1 экспрессирует один основной и два минорных транскрипта мРНК, различающихся только длиной 5′-нетранслируемых лидерных последовательностей, предшествующих общей открытой рамке считывания (ORF) (Sunderland et al. 2006). Контроль продукции изоформы AtLIG1 и внутриклеточное нацеливание зависит от механизмов, контролирующих выбор сайта инициации трансляции в пределах ORF AtLIG1.Трансляция транскриптов мРНК AtLIG1 с первого стартового кодона в рамке считывания дает изоформу AtLIG1, которая нацелена исключительно на митохондрии. Инициация трансляции со второго стартового кодона в кадре дает изоформу AtLIG1, нацеленную только на ядро. Длина 5′-UTR и, что более важно, нуклеотидный контекст вокруг альтернативных стартовых кодонов в транскриптах AtLIG1 влияют на инициацию трансляции для обеспечения сбалансированного синтеза ядерных и митохондриальных изоформ AtLIG1 через контекстно-зависимый механизм сканирования дырявых рибосом (Sunderland et al. .2006 г.). Вместо того, чтобы иметь отдельные гены для кодирования различных изоформ белков ДНК-лигазы, специфичных для компартментов, эукариотические виды, от дрожжей до человека, по-видимому, используют эволюционно законсервированный механизм, который основан на выборе стартовых кодонов инициации трансляции в рамке считывания внутри рамки считывания транскрипта мРНК лигазы. для регулирования синтеза соответствующей изоформы ДНК-лигазы, предназначенной для ядра или митохондрий. Поскольку контроль субклеточной локализации белков и их взаимодействия со специфическими белками-партнерами in vivo имеют решающее значение для физиологии клетки, важность механизмов, контролирующих производство различных форм белка и их внутриклеточную топологию, трудно переоценить (Sunderland et al. 2006 г.).
Ионные каналы состоят из трансмембранных белков, которые позволяют ионам проникать с высокой скоростью. Каналы K + (Kv) с затвором напряжения незаменимы для электрической возбудимости нервных и мышечных волокон (Finol-Urdaneta et al. 2006). N-концевая область Kv-каналов играет важную регуляторную роль, включая кинетику инактивации, распознавание субъединиц и окислительно-восстановительную модуляцию токов, протекающих по этим каналам. Таким образом, различия между N-концевыми участками Kv каналов могут приводить к важным функциональным различиям между молекулярными формами Kv каналов.Каналы Kv 1.7 от сердечной мышцы мыши кодируются геном Kcna7 . Его транскрипт мРНК имеет два предполагаемых сайта инициации трансляции, которые генерируют изоформы двух каналов, mKv1.7L (489 аминокислотных остатков) и mKv1.7S (457 аминокислотных остатков), с различными электрофизиологическими характеристиками. Наличие обеих изоформ очень важно для нормального функционирования миокарда, поскольку отсутствие одной из них приводит к различным типам сердечной патологии (Finol-Urdaneta et al. 2006).
Инсулин-деградирующий фермент (IDE) экспрессируется как в чувствительных к инсулину, так и в нечувствительных к инсулину тканях.Внутри клетки IDE нацелена преимущественно на цитоплазму, однако в некоторых клетках она представлена также в нескольких субклеточных компартментах. Механизмы, ответственные за нацеливание IDE на разные клеточные компартменты, не совсем понятны. Изучая этот вопрос, Leissring et al. (Leissring et al. 2004) показали, что альтернативная трансляция, начинающаяся с инициирующего кодона в рамке считывания, расположенного перед каноническим стартовым сайтом, генерирует более раннюю нераспознанную изоформу деградирующего инсулин фермента, которая в отличие от изоформы, находящейся в цитоплазме, нацелена на митохондрии.Этот пример показывает, что N-концевое удлинение, достигаемое за счет использования альтернативного инициирующего кодона, может продуцировать новую изоформу белка, обладающую новой функцией.
Эти и многие другие примеры (Porras et al. 2006; Sass et al. 2001) демонстрируют силу механизмов просачивающегося сканирования в генерации множества функционально различных изоформ белков из одного транскрипта мРНК.
b) Шунтирование рибосом
Следующим механизмом трансляции, который также влияет на разнообразие белковых продуктов, кодируемых одним геном, является шунтирование рибосом.Модель сканирования постулирует, что когда сканирующая 40S рибосомная субъединица встречает шпильку в 5 ‘UTR, она не пропускает петлю, а раскручивает ее (Kozak, 1980; Lopez-Lastra et al. 2005). Тем не менее, есть некоторые случаи, когда сканирующая 40S рибосомная субъединица встречает структуры, присутствующие в 5’UTR, и пропускает или шунтирует большой сегмент, минуя промежуточные сегменты, включая кодоны AUG и сильные вторичные структуры, которые обычно блокируют процесс сканирования (). Селективная инициация трансляции в инфицированных аденовирусом клетках при тепловом шоке является примером механизма прыжка рибосом (Yueh and Schneider, 1996).
c) Завершение-инициирование
Третий неклассический механизм инициирования, зависящий от ограничения, как уже упоминалось выше, — это завершение-инициирование. В механизме повторной инициации вторая ORF, расположенная в той же мРНК, может транслироваться без того, чтобы субъединица 40S рибосомы отделялась от мРНК после достижения первого стоп-кодона ORF (). Строго говоря, этот механизм больше связан со стадией элонгации, чем с инициирующей, хотя для продолжения трансляции необходим шаг повторной инициации в последующем триплете AUG.
Исследование механизма повторной инициации терминации для регуляции трансляции белков наблюдается в различных клетках и вирусах. Напр., Стресс-индуцированное фосфорилирование eIF2α увеличивает трансляцию ATF4, активатора программы интегрированного стрессового ответа (ISR), по механизму повторной инициации (Lu et al. 2004). У мРНК ATF4 есть две консервативные восходящие ORF (uORF), uORF1 и uORF2. Сканирующие рибосомы инициируют трансляцию в обеих uORF. После трансляции uORF сканирующая рибосома эффективно повторно инициирует трансляцию в последующих AUG.В нестрессированных клетках низкие уровни фосфорилирования eIF2α способствуют ранней активации таких повторно инициирующих рибосом, направляя их к ингибирующей uORF2, что препятствует последующей трансляции ATF4 и подавляет ISR. В стрессированных клетках высокие уровни фосфорилирования eIF2α задерживают емкость рибосом и способствуют повторной инициации на ATF4 по сравнению с ингибирующим uORF2 (Lu et al. 2004). Аналогичный механизм действует в регулируемой трансляции белка Gsn4 у дрожжей (Hinnebusch and Natarajan, 2002).
Трансляция полимеразы вируса гепатита человека — еще один пример, демонстрирующий участие различных механизмов трансляции, участвующих в экспрессии полицистронной мРНК.Вирус гепатита В человека (HBV) имеет компактный геном, кодирующий четыре основных перекрывающихся кодирующих области: ядро, полимеразу, поверхность и X. Кодону инициации полимеразы предшествует частично перекрывающееся ядро и четыре или более вышестоящих кодонов инициации. Несколько механизмов используются для включения синтеза белка полимеразы, включая сканирование с утечкой и повторную инициацию рибосом. Chen et al. (Chen et al. 2005a) показали, что основная открытая рамка считывания (CO uORF), высококонсервативная для всех подтипов HBV, играет ингибирующую роль в экспрессии нижестоящей от основного сайта инициации, но также стимулирует повторную инициацию в начале полимеразы, когда она находится в оптимальный контекст.Таким образом, CO uORF является определяющим фактором в уравновешивании синтеза белков ядра и полимеразы.
2. Кеп-независимая инициация, определяемая IRES
Описанные примеры демонстрируют возможности кэп-зависимых механизмов инициации для генерации множества изоформ белка, обладающих новыми функциями. Однако возможности процесса трансляции увеличивать и диверсифицировать потенциалы, встроенные в мРНК, не ограничиваются только кэп-зависимыми механизмами, поскольку независимые от кэпа механизмы трансляции действуют также в клетках.Cap-независимая трансляция () обеспечивается внутренними сайтами входа в рибосомы, IRES (Holcik et al. 2000). Они обычно состоят из структурированных областей в 5′-нетранслируемой области и первоначально были идентифицированы у вирусов, но с тех пор были обнаружены в мРНК эукариот и, как полагают, составляют основную форму регуляции синтеза белка в клетках млекопитающих (Candeias et al. 2006). ; Holcik et al. 2000). Как правило, между отдельными IRES нет значительного структурного сходства.В настоящее время IRES определяются исключительно функциональными критериями и еще не могут быть предсказаны по присутствию характерных последовательностей РНК или структурных мотивов (Lopez-Lastra et al. 2005). Изучение доступного программного обеспечения для прогнозирования структуры РНК и программ поиска мотивов РНК показывает, что, хотя эти программы являются полезными инструментами для точной настройки эмпирически определенной вторичной структуры РНК, точность предсказания вторичной структуры de novo больших молекул РНК и последующей идентификации новых Использование элементов IRES с помощью вычислительных подходов по-прежнему невозможно (Baird et al.2006 г.).
a) Смешанная (или альтернативная) инициация
Важно, чтобы некоторые клеточные мРНК, хотя и кэпированные, включая те, которые кодируют факторы инициации трансляции, факторы транскрипции, онкогены, факторы роста, продукты гомеотических генов и белки выживания, также содержали элементы IRES. в их 5’UTR-последовательностях, которые могут позволить им транслироваться в условиях, когда кэп-зависимый синтез белков нарушен (Lopez-Lastra et al. 2005). Более того, в некоторых случаях молекула мРНК содержит как кэп-зависимый стартовый кодон, так и элемент IRES, и в различных клеточных условиях может управлять синтезом различных изоформ одного белка с использованием кэп-зависимых или IRES-опосредованных механизмов трансляции ().Например, экспрессия двух изоформ p53, полной длины (FLp53) и усеченной на N-конце (p53 / 47), контролируется альтернативными кэп-зависимыми и IRES-опосредованными механизмами инициации, регулирующими трансляцию мРНК p53 (Candeias et al. 2006 г.). В отличие от FLp53, изоформа p53 / 47 не содержит большей части N-концевого трансактивирующего домена p53 (aa 1-40) или сайта связывания Mdm2 (aa 17-23), демонстрирующего разные функциональные параметры. P53 / 47 может образовывать комплексы с FLp53, тем самым изменяя его стабильность и способность индуцировать активацию других p53-зависимых генов.Изменения в синтезе FLp53 или p53 / 47 регулируются посредством различных индуцированных клеточным стрессом путей, действующих через отдельные участки мРНК p53. Функциональные различия двух изоформ проявляются также в том, что для некоторых цитотоксических агентов требуется присутствие FLp53 для индукции апоптоза, тогда как для других достаточно p53 / 47. Это указывает на то, что, укрывая альтернативные сайты инициации трансляции, мРНК р53 дает начало различным уровням изоформ р53, которые помогают управлять биологическим результатом активации р53 в ответ на различные типы клеточного стресса (Candeias et al.2006 г.).
Другим примером, показывающим роль альтернативной инициации трансляции в синтезе изоформ белков, обладающих различными функциями, является фактор роста эндотелия сосудов (VEGF). Необычно длинный (1038 нуклеотидов) и структурированный 5′-нетранслируемый участок (UTR) мРНК VEGF содержит два независимых IRES (IRES A и IRES B). В человеческой последовательности четыре потенциальных кодона инициации трансляции CUG расположены между этими IRES и находятся в рамке с классическим стартовым кодоном AUG.VEGF IRES A, расположенный в пределах 300 нуклеотидов выше кодона AUG, управляет инициацией кэп-независимой трансляции, которая, как было показано, позволяет синтез VEGF в условиях гипоксии (Huez et al. 1998; Sass et al. 2001; Stein et al. 1998) ). Было показано, что IRES B, расположенный в первой половине 5’UTR между нуклеотидами 379 и 483, контролирует инициацию трансляции новой изоформы VEGF (L-VEGF), которая удлинена на N-конце на 206 аминокислот по сравнению с классической изоформой VEGF. Форма, инициированная AUG.Инициация трансляции этой изоформы происходит по первому из кодонов CUG. О различных функциях двух изоформ VEGF может свидетельствовать различное нацеливание изоформ, инициированных AUG и CUD: в то время как классическая изоформа VEGF, инициированная AUG, является секретируемым белком, изоформа L-VEGF, инициированная CUG, демонстрирует внутриклеточное нацеливание. Кроме того, L-VEGF подвергается посттрансляционному частичному расщеплению на два фрагмента, а именно на 206-а.о. N-концевой фрагмент, который после расщепления остается в клетке, и C-концевой фрагмент, который равен изоформе, инициированной AUG, по размеру и способности секретироваться из клетки (Huez et al.2001).
B. Стадия элонгации
Таким образом, на стадии инициации процесса трансляции аппарат трансляции имеет многочисленные возможности для материальной (молекулярной) реализации скрытых в мРНК информационных потенциалов. На стадии элонгации другие механизмы также могут влиять на структурное и функциональное разнообразие синтезируемых изоформ белков. В отличие от стадий инициации и завершения трансляции, механизмы, используемые во время удлинения трансляции, были в высокой степени консервативными и менее изменчивыми в трех царствах жизни (Kapp and Lorsch, 2004).Как отмечают Namy et al. (Namy et al. 2004), основными механизмами, с помощью которых машины трансляции могут обнаруживать скрытую генетическую информацию на стадии элонгации, являются смещение рамки () и, в меньшей степени, прыжки ().
1. Сдвиг кадра
Предсказание белка (белков), кодируемого данной ДНК, затруднено из-за отсутствия знаний о сигналах, встроенных в геном, которые участвуют в трансляции генетической информации. Было показано, что во многих генах стандартные правила декодирования, постулируемые догмой универсального генетического кода, подрываются путем «перекодирования» сигналов, обнаруженных в ДНК, которые способствуют альтернативным событиям декодирования, таким как запрограммированный сдвиг рамки считывания, скачкообразный переход или перераспределение терминирующего кодона (Аткинс). и другие.2001; Гестеланд и Аткинс, 1996; Gesteland et al. 1992).
Как отмечено Namy et al. (Namy et al. 2004) «естественные ошибки сдвига рамки возникают очень часто, но запрограммированные рибосомные сигналы сдвига рамки чрезвычайно увеличивают вероятность проскальзывания тРНК, иногда до такой степени, что до 50% рибосом меняют рамку». Анализ механизмов смены кадров выходит за рамки данного обзора. Они были критически изучены и описаны в нескольких прекрасных публикациях (Atkins et al., 2001; Brierley, Pennell, 2001; Plant et al.2003; Stahl et al. 2001). В принципе, смещение кадра запускается двумя элементами: скользким движением и последовательностью стимулятора. Скользкая последовательность — это область мРНК, в которой движение или смещение тРНК предпочтительнее, тогда как стимулятор — это последовательность мРНК, которая усиливает смещение фрейма за счет индукции рибосомной паузы (Namy et al. 2004).
Различные виды смены кадра описаны как у прокариот, так и у эукариот. Например, запрограммированный сдвиг рамки наблюдался при трансляции прокариотической ДНК-полимеразы III, α-фукозидазы и фактора высвобождения 2 (RF2), а также эукариотических белков, таких как антизим-1 (AZ1), IL-10, ABP140 и т. Д. [Цит. Namy et al.2004)]. Биологическая целесообразность и функции такого сдвига рамки в каждом случае разные, но всегда оправданы. Например, в случае RF2 сдвиг кадра перекодирования представляет собой механизм саморегулирования, контролирующий количество RF2. При высоких уровнях RF2 конкуренция между завершением и сдвигом кадра смещается в пользу завершения, что приводит к снижению концентрации RF2 в клетке. Когда уровень RF2 начинает снижаться, начинает преобладать сдвиг кадра, повышая уровень RF2 (Adamski et al.1993).
Антизим-1 (AZ1) — еще один хорошо изученный пример белка, полная экспрессия которого регулируется запрограммированным сдвигом рамки. AZ1 — это белок, который негативно регулирует синтез полиамина, ингибируя ключевой синтетический фермент орнитиндекарбоксилазу и направляя его на деградацию протеасомой 26S (Heller et al. 1976; Murakami et al. 1992). Как и RF2, экспрессия полноразмерного белка AZ1 зависит от сдвига рамки считывания, который, в свою очередь, связан с механизмом обратной связи, который регулирует уровни полиамина в клетках млекопитающих.Для AZ1, когда клеточные уровни полиаминов низкие, продуцируется полипептид, соответствующий аминокислотным остаткам 1–68, и трансляция заканчивается на следующем стоп-кодоне UGA (Matsufuji et al. 1995). Когда клеточные уровни полиаминов высоки, сдвиг рамки считывания на +1 происходит после 68-го кодона, что приводит к полноразмерному антизимному белку, содержащему 227 аминокислотных остатков (1-227 а.о.) (Ichiba et al. 1994). Сдвиг рамки используется при экспрессии всех известных антизимов у разных видов и многих других белков.Согласование этого механизма на протяжении всей эволюции подчеркивает решающую роль сдвига рамки считывания в регуляции экспрессии генов.
Следует отметить, что сдвиг кадров лишь частично определяет многофункциональность AZ1. Дифференциальное использование альтернативных сайтов инициации также оказывает свое влияние на это явление. МРНК AZ1 содержит два инициирующих кодона AUG, разделенных 32 кодонами. Оба триплета AUG используются в качестве сайтов начала трансляции (Matsufuji et al. 1995; Rom and Kahana, 1994).Было показано, что N-концевой регион AZ1 содержит сигналы ядерного экспорта и нацеливания на митохондрии (Gandre et al. 2003; Gritli-Linde et al. 2001; Murai et al. 2003). В зависимости от того, какой кодон AUG используется для трансляции, полученная изоформа AZ1 нацелена на ядро, митохондрии или цитоплазму (Murai et al. 2003). Многофункциональность антизима подтверждается не только направлением его изоформ в различные компартменты клетки и использованием в качестве ингибитора орнитиндекарбоксилазы, но также недавним открытием того, что AZ1 функционирует как положительный транскрипционный регулятор генов оперона atoDAEB у E.coli (Филиппоу и др., 2007).
Мы описали здесь только два белка, синтез которых направлен путем сдвига рамки считывания. Многие другие примеры, в которых этот механизм используется для развития разнообразия белков, можно найти в очень информативном обзоре, написанном Namy et al. (Нами и др., 2004).
2. Перескок (обход трансляции)
Необычным событием сдвига рамки считывания, которое также диверсифицирует структуры и функции белка, является «скачкообразный переход», когда переход от кадра 0 к другому кадру происходит путем обхода трансляции расширенной области последовательности мРНК, а не проскальзыванием одного нуклеотида, как было описано для большинства примеров сдвига рамки.Когда был открыт генетический код, его перевод казался жестким. Однако позже было обнаружено, что чтение генетического текста (кода) довольно гибкое, и было описано несколько альтернатив его выражения (Groisman and Engelberg-Kulka, 1995).
Трансляционный обход объединяет информацию, обнаруженную в двух разрозненных открытых рамках считывания, в единую полипептидную цепь. Основной механизм основан на свойствах декодирования пептидил-переносящей РНК (тРНК) и включает три стадии: взлет, , в котором комплекс пептидил-тРНК / мРНК в Р-сайте рибосомы диссоциирует; сканирование, , при котором пептидил-тРНК зондирует мРНК, скользящую через центр декодирования; и посадки, , в котором пептидил-тРНК восстанавливается кодоном, с которым она может образовывать стабильное взаимодействие () (Herr et al.2000а). Лучше всего охарактеризованными примерами этого явления являются ген 60 Т4 и ген plaA (ген адгезии Prevotella loescheii ). В гене 60 Т4 комплекс сигналов стимулирует обход 50 нуклеотидов между двумя открытыми рамками считывания (Herr et al. 2000b). В plaA трансляция его мРНК требует обхода 29-нуклеотидной кодирующей бреши (Manch-Citron et al. 1999).
Herr et al. (Herr et al. 2000a) отметили, что хотя известно несколько примеров генов, которые полагаются на обход трансляции для пары ORF, рибосомы, по-видимому, обладают врожденной способностью обхода.Это говорит о том, что стратегия обхода трансляции может быть более распространенной, чем это принято в настоящее время.
C. Стадия терминации
Прекращение синтеза белка включает декодирование стоп-сигнала через взаимодействие между рибосомой, мРНК и факторами высвобождения (RF), что способствует гидролитическому высвобождению растущей полипептидной цепи из пептидилтрансферазного центра рибосома (Кридж и др., 2006). Стоп-кодоны используются для подачи сигнала рибосоме о прекращении декодирования матрицы мРНК.Открытые рамки считывания в мРНК оканчиваются внутрикадровым любым из трех стоповых или бессмысленных кодонов (UAA, UAG или UGA), которые распознаются во время трансляции путем связывания RF с сайтом рибосомы A (von der Haar and Tuite, 2007; Уильямс и др. 2004).
1. Считывание стоп-кодонов
Прекращение трансляции обычно является высокоэффективным процессом. Было показано, что стоп-кодоны обычно подавляются с частотой всего 0,001–0,1% (Loftfield and Vanderjagt, 1972; Stansfield et al.1998). Когда стоп-кодон достигает A-сайта транслирующейся рибосомы, могут происходить два конкурирующих события: завершение и считывание стоп-кодона (или бессмысленное подавление). Прекращение трансляции приводит к гидролизу последней связи пептидил-тРНК и высвобождению завершенного незрелого полипептида. Альтернативно, чтение, при котором стоп-кодон ошибочно декодируется супрессором или близкородственной т-РНК, приводит к трансляции мимо стоп-кодона и продукции белка с С-концевым удлинением ().Относительная частота терминации по сравнению с прочтением определяется такими параметрами, как контексты нуклеотидов стоп-кодона выше и ниже, активности факторов высвобождения терминальной полипептидной цепи и количество супрессоров, близких к родственным т-РНК. Все эти параметры модулируют баланс реакций терминации и прочтения в зависимости от типа клетки (Bonetti et al. 1995; Fearon et al. 1994; Janzen and Geballe, 2004; Keeling et al. 2004; Namy et al. 2002). . Willams et al.(Williams et al. 2004) подчеркивают, что «потенциал даже короткого C-концевого удлинения (пептидных меток) для изменения клеточной локализации или активности белка велик. Добавление даже одной аминокислоты может завершить частичный сигнал нацеливания, уже присутствующий на С-конце белка. Все расширения белка, изменяющие нацеливание, потенциально могут действовать как генетически доминантные события, связанные с усилением функции. Таким образом, несмотря на в целом низкую эффективность событий считывания стоп-кодонов, такие события вполне могут иметь фенотипические последствия для клетки ».
Существует все больше примеров считывания стоп-кодонов как в вирусных, так и в клеточных системах, и во многих случаях это считывание имеет физиологические последствия для клетки. Стало очевидным, что разные клетки и организмы не только принимают стоп-кодон в качестве основного сигнала терминации, но также используют его для обеспечения гибкости этих клеток и организмов для физиологической реакции на различные изменчивые условия (Namy et al., 2002; Steneberg and Samakovlis, 2001). ; Тейт и др.1999; Yoshinaka et al. 1985). Альтернативное неканоническое использование стоп-кодонов позволяет продуцировать несколько полипептидов из одной открытой рамки считывания, тем самым увеличивая структурное и функциональное разнообразие продуктов одного гена (Doronina and Brown, 2006). Классическая иллюстрация к этому утверждению — селенопротеины.
Описано более 15 селенопротеинов животных. Многие селенопротеины, такие как глутати-онпероксидаза (Sukenaga et al., 1987), йодтиронин-5′-дейодиназа I типа (5’DI) (Berry et al.1991b) и тиоредоксинредуктаза (Gasdaska et al. 1995) являются ферментами, которые катализируют окислительно-восстановительную реакцию. Все селенопротеины содержат селеноцистеин (Sec). Включение селеноцистеина определяется кодоном UGA в открытой рамке считывания мРНК, который сопровождается элементом «последовательности вставки селеноцистеина» (SECIS) в 3′-UTR (Berry et al. 1991a; Goto et al. 2001). Однако в определенных условиях этот кодон UGA может также служить стоп-кодоном. Конкуренция между UGA в качестве стоп-сигнала и кода для Sec наблюдалась во время экспрессии гена дейодиназы.Как продукт терминации (14 кДа), так и полный белок йодиназы (28 кДа) могут быть обнаружены в клетках (McCaughan et al. 1995). Относительные количества этих продуктов могут варьироваться в зависимости от типа клетки и системы экспрессии, отражая конкуренцию между двумя механизмами декодирования UGA (Steneberg and Samakovlis, 2001).
Селенопротеин P представляет еще более сильную иллюстрацию конкуренции между UGA как стоп-кодоном и UGA как триплетом, который определяет Sec-включения. Селенопротеин P представляет собой внеклеточный гликопротеин, который, как предполагается, служит для защиты от окислителя (Burk et al.1995). Его мРНК содержит 10 кодонов UGA в открытой рамке считывания, оканчивающейся UAA, что подразумевает наличие 10 селеноцистеинов в первичной структуре белка (Hill et al. 1991). Полноразмерный селенопротеин P и три меньшие изоформы, которые имеют идентичные N-концы, но разное сродство к гепарин-сефарозе, были обнаружены в плазме крыс (Chittum et al. 1996). Путем масс-спектрометрического анализа пептидов было доказано, что полноразмерная изоформа содержит предсказанные аминокислотные остатки, включая ее С-конец и все 10 селеноцистеинов.Укороченные изоформы заканчивались там, где, как предполагалось, располагались второй, третий и седьмой остатки селеноцистеина (Ma et al. 2002). Это предполагает, что все изоформы возникают из одной и той же мРНК и что UGA, которые определяют второй, третий и седьмой селеноцистеины в полноразмерном селенопротеине P, могут альтернативно служить для прекращения трансляции, продуцируя более короткие изоформы. Как отмечает Ma et al. (Ma et al. 2002) указали, что «остается теоретической возможность того, что более короткие изоформы продуцируются из полноразмерного белка путем протеолиза».Однако протеаза, которая могла бы расщеплять белок по специфическому остатку селеноцистеина, неизвестна, и это исключает возможность протеолитического расщепления полноразмерной изоформы по специфическим Sec-сайтам.
Ген Drosophila kelch является другим примером, показывающим, что трансляционный контроль экспрессии гена может быть реализован посредством подавления и считывания стоп-кодонов. Робинсон и Кули (Robinson and Cooley, 1997) продемонстрировали, что ген kelch продуцирует единственный транскрипт со стоп-кодоном UGA, разделяющим две открытые рамки считывания (ORF1 и ORF2).Из транскрипта экспрессируются 76 кДа ORF1 и 160 кДа полноразмерные (ORF1 + ORF2) белки. Экспрессия этих двух белков регулируется тканеспецифическим образом, в результате чего соотношение двух изоформ варьируется в разных тканях (Robinson and Cooley, 1997).
Важную роль «протекающего» стоп-кодона и механизма считывания для физиологии клетки можно также проиллюстрировать на примере дрожжевого гена PDE2 , который кодирует высокоаффинную фосфодиэстеразу цАМФ. Путем синтеза цАМФ-фосфодиэстеразы этот ген может регулировать внутриклеточную концентрацию цАМФ и, таким образом, защищать клетку от внеклеточного цАМФ (Wilson et al.1993; Уилсон и Тэтчелл, 1988). Когда трансляция мРНК PDE2 завершается на «дырявом» стоп-кодоне, синтезируется белок pde2 (активная фосфодиэстераза), что приводит к низкому уровню внутриклеточного цАМФ и повышенной чувствительности к стрессу. Напротив, когда включается механизм считывания стоп-кодонов, происходит удлинение С-конца 21 аминокислоты, что приводит к дестабилизации белка pde2, что, в свою очередь, приводит к высокому уровню цАМФ в клетке и устойчивости к стрессовой среде (Namy et al. .2002; фон дер Хаар и Туите, 2007). Таким образом, PDE2 является примером продукта отдельного гена в Saccharomyces cerevisiae , где изменения в считывании стоп-кодона напрямую связаны с фенотипическими вариациями, которые имеют решающее значение для физиологии клетки.
Когда обсуждается физиологическая роль канонических стоп-кодонов, протекающих стоп-кодонов и механизмов чтения, основное различие между нормальной функцией стоп-кодона терминации и аномальной ролью преждевременных стоп-кодонов (ов), которые могут появиться в мРНК в результате мутаций или альтернативный сплайсинг с загадочными сайтами сплайсинга следует подчеркнуть (Frischmeyer and Dietz, 1999; Lewis et al.2003 г.). Нуклеотидный контекст, окружающий классический стоп-кодон, определяется генетически и способствует прекращению трансляции синтезированного природного белка. Контексты восходящего и нисходящего потока сигнала остановки утечки также генетически детерминированы для стимулирования активности считывания, что приводит к появлению новой изоформы белка, необходимой для выполнения определенной физиологической функции. Таким образом, экспрессия терминированных или удлиненных белков в этих случаях представляет собой реализацию определенных этапов нормальной клеточной программы.Случайное появление преждевременного терминирующего кодона (PTC) или бессмысленного кодона в ORF мРНК не коррелирует с унаследованным окружающим нуклеотидным контекстом и, следовательно, не может регулироваться физиологическими механизмами трансляции (Lewis et al. 2003).
Транскрипты, несущие бессмысленные кодоны, обычно кодируют усеченные на С-конце белки, которые могут обладать доминантно-отрицательными или пагубными эффектами усиления функции. Обычно PTC запускают быструю деградацию мРНК с помощью механизма нонсенс-опосредованного распада мРНК (NMD), который обнаруживает и разрушает транскрипты, несущие нонсенс-кодоны (Frischmeyer and Dietz, 1999; Wilkinson, 2005).Эта негативная реакция является важным механизмом контроля качества. Его функция — защита клеток от аномальной функции мРНК (Behm-Ansmant et al. 2007; Isken and Maquat, 2007). Удивительно, но недавние исследования убедительно доказали, что компоненты аппарата NMD (белки UPF1, UPF2, UPF3, RNP21, Y14 и MAGOH) также способствуют трансляции нормальных мРНК в клетках млекопитающих (Nott et al. 2004; Wiegand et al. 2003; Wilkinson, 2005). ). Было показано, что функция белков NMD зависит от их местоположения.Если факторы NMD расположены в пределах ORF, они стимулируют трансляцию и контролируют завершение трансляции с высокой точностью. Если они расположены ниже ORF, они вызывают распад РНК (Lykke-Andersen et al. 2001; Maderazo et al. 2000; Wang et al. 2001; Wiegand et al. 2003; Wilkinson, 2005). Таким образом, белки NMD также демонстрируют многофункциональность, участвуя в двух противоположных процессах. С одной стороны, участвуя в NMD, они разрушают мРНК и снижают уровень синтеза белка. С другой стороны, связываясь с мРНК в пределах ORF, они стимулируют синтез белков и даже исправляют естественные ошибки (Wilkinson, 2005).
Описанные примеры не только документируют существование нескольких эволюционно консервативных механизмов трансляции, предназначенных для продукции множества и функционально различных изоформ одного продукта гена, но также показывают, что белки, участвующие в этих механизмах, являются многофункциональными молекулами, которые одновременно могут участвовать в противоположных процессах, ведущих к для уменьшения и увеличения синтеза белка. Эти механизмы оказывают существенное влияние на преобразование компактной генетической информации в различные белковые фрагменты.
Посттрансляционные модификации
Хотя аминокислотная структура белка определяется геном и мРНК, кодирующими белок, его окончательная функциональная структура достигается за счет многочисленных посттранскляционных модификаций. Большинство всех белков подвергаются ко- и / или посттрансляционным модификациям (Rehfeld and Goetze, 2003). Знание этих модификаций чрезвычайно важно, поскольку они могут изменять физические и химические свойства, распределение конформации, укладку, стабильность, активность и, следовательно, функцию белков.Предшественник белка может подвергаться широкому спектру протеолитических расщеплений, обрезков N- и C-концов и дериватизации аминокислот в клетках, которые экспрессируют белок. Иногда один и тот же предшественник по-разному процессируется в разных типах клеток (Rehfeld and Goetze, 2003). Согласно Rehfeld и Goetze (Rehfeld and Goetze, 2003) все посттрансляционные модификации можно разделить на три группы [цит. из (Rehfeld and Goetze, 2003)].
Таблица 1
Посттрансляционные модификации.
Эндопротеолитические расщепления:
Двухосновные сайты
Одноосновные сайты
Post (поли) Clu / Asp сайты
-9000 сайтов
-9000 Post
Экзопротеолитические тримминги:
Карбоксамидирование
Глутаминилциклирование
N-концевое дипептидильное расщепление
4
N-концевое дипептидильное расщепление 1
Дисульфидные мостики
Гамма-карбоксилирование
Гликозилирование
Изомеризации
Фосфорилирование
pes посттрансляционной модификации, его нельзя рассматривать как полный список возможных модификаций.Например, он не содержит таких модификаций, как пальмитоилирование, убиквитилирование, метилирование, сворачивание белка и сплайсинг белка, которые, по-видимому, являются мощным механизмом разнообразия белков (Draper et al. 2007; Hanada et al. 2004; Mostaqul Huq et al. 2008). ; Наградова, 2004; Перлер, 2005; Шартнер и др. 2007). Ограничение объема обзора не позволяет обсудить все возможные посттрансляционные модификации, изображенные в. Поэтому мы сосредоточимся только на некоторых из них — фосфорилировании, гликозилировании, пальмитоилировании, сворачивании белков и сплайсинге белков.
1. Фосфорилирование белка
Фосфорилирование белка — один из наиболее распространенных типов посттрансляционной модификации белка, который позволяет клетке контролировать различные клеточные процессы, включая передачу сигнала, механизмы транскрипции и трансляции, метаболизм, рост, деление, дифференцировку. , подвижность, транспорт органелл, мембранный транспорт, иммунитет, обучение и память (Manning et al. 2002a; Manning et al. 2002b). В эукариотических клетках наибольшее фосфорилирование происходит по остаткам Ser, Thr и Tyr (Ubersax and Ferrell, 2007).Состояние фосфорилирования фосфопротеинов контролируется активностью протеинкиназ и фосфатаз (Urner and Sakkas, 2003). Согласно расчетам, проведенным Уберсаксом и Ферреллом (Ubersax and Ferrell, 2007), типичная протеинкиназа должна распознавать от одного до нескольких сотен истинных сайтов фосфорилирования на фоне примерно 700000 потенциально фосфорилируемых остатков в типичной эукариотической клетке. Разработано множество механизмов, которые способствуют этой исключительной специфичности, включая структуру каталитического сайта, локальные и удаленные взаимодействия между киназой и субстратом, образование комплексов с каркасными и адапторными белками, которые пространственно регулируют киназу и т. Д. (Ubersax and Ferrell, 2007 ).
Хотя разные киназы демонстрируют свои индивидуальные особенности и особенности в процессах фосфорилирования, основными результатами этой посттрансляционной модификации являются структурные и функциональные изменения белка-субстрата. Это утверждение можно проиллюстрировать несколькими примерами.
Было обнаружено, что многие ядерные рецепторы модифицируются путем фосфорилирования (Rochette-Egly, 2003). Рецептор яичек 4 (TR4) является сиротским членом надсемейства ядерных рецепторов. В отсутствие специфических лигандов активность TR4 может модулироваться посредством фосфорилирования его домена AF-1, опосредованного митоген-активируемой протеинкиназой (MAPK) (функция активации 1).MAPK-опосредованное фосфорилирование домена AF-1 делает TR4 репрессором, тогда как дефосфорилирование домена AF-1 делает TR4 активатором (Huq et al. 2006). Обратимое фосфорилирование белков, как и двунаправленная регуляция TR4, как известно, контролирует широкий спектр биологических активностей (Chen et al. 2005b; Collins et al. 2004; Kim et al. 2003; Koritschoner et al. 2001; Shyr et al. 2002). ; Tanabe et al.2002).
Другой пример также может показать силу посттрансляционного фосфорилирования в определении функции белка.Фосфорилирование, опосредованное глюкозой, превращает фактор транскрипции Rgt1 из репрессора в активатор (Mosley et al. 2003). В дрожжах Saccharomyces cerevisiae глюкоза индуцирует экспрессию транспортируемых гексозой генов / HXT / путем модуляции активности фактора транскрипции Rgt1, который функционирует как репрессор, когда глюкоза отсутствует. Однако в присутствии высокой концентрации глюкозы Rgt1 превращается из репрессора в активатор путем фосфорилирования, опосредованного глюкозой.Активатор Rgt1 необходим для максимальной индукции экспрессии гена HXT1 (Mosley et al. 2003).
Было показано, что несколько факторов транскрипции, а также другие белки изменяют свою субклеточную локализацию в ответ на внешние стимулы (De Vit et al. 1997; Komeili and O’Shea, 1999). Дрожжевой белок Mig1 фосфорилируется киназой Snf1 в отсутствие глюкозы и задерживается в цитоплазме. Высокие концентрации глюкозы вызывают дефосфорилирование Mig1 и транслокацию в ядро (Komeili and O’Shea, 1999; Treitel et al.1998). Субклеточная локализация другого дрожжевого белка, Pho4, регулируется в ответ на другой агент, фосфат. При высоких концентрациях фосфата Pho4 фосфорилируется и экспортируется в цитоплазму, тогда как голодание по фосфату вызывает дефосфорилирование Pho4 и перемещение в ядро (Komeili and O’Shea, 1999).
Эти примеры ясно демонстрируют, что определенные функциональные потенциалы, закодированные в первичной структуре белка с помощью определенной топологии остатков Ser, Thr и Tyr, могут быть обнаружены и реализованы посредством определенных событий фосфорилирования.
2. Гликозилирование
Гликозилирование — еще одна широко наблюдаемая посттрансляционная модификация белков. Белки могут быть гликозилированы на боковых цепях определенных аминокислот, и эти модификации обозначаются как N- и O-гликозилирование (Medzihradszky, 2002). N-гликозилированные частицы модифицируются по остаткам Asn. О-гликозилирование происходит по остаткам Ser или Thr. Следует отметить, что характеристика обоих типов гликозилирования осложняется тем фактом, что в некоторых случаях одни и те же аминокислоты в популяции белковых молекул могут быть дериватизированы с набором различных углеводных структур, в то время как в других случаях они могут оставаться неизмененными.Эта сайт-специфическая гетерогенность может варьироваться в зависимости от вида, ткани и может зависеть от физиологических изменений (Medzihradszky, 2002). Таким образом, один и тот же белок может быть гликозилирован в одном типе клеток и немодифицирован в другом типе клеток, и разница в статусе гликозилирования будет отражаться разными функциями данного белка в разных клетках (Bloom et al., 1996; Hironaka et al. 1993). Аналогичная ситуация может наблюдаться даже в одних и тех же клетках, однако в разных физиологических условиях (Nemansky et al.1998).
Целью гликозилирования, как и любой другой посттрансляционной модификации, является выявление функциональных активностей, скрытых в структуре белка. При добавлении сахаров к молекуле полипептида гликозилирование изменяет ее структуру таким образом, что могут возникать новые функции. Например, Миранда и др. (Miranda et al. 2007) показали, что нейтрализующие свойства ВИЧ-специфического антитела F240 резко изменяются в результате гликозилирования. Это Ab при получении в гибридоме не является нейтрализующим.Однако Ab F240 IgG1, экспрессируемые в клетках CHO, приобретали сильную нейтрализующую активность без изменения иммунореактивности. Секвенирование мРНК F240, продуцируемой в родительской гибридоме и клетках СНО, выявило идентичные последовательности, предполагая, что приобретенная нейтрализация является результатом клеточно-специфичных посттрансляционных модификаций. Среди различных типов посттрансляционных модификаций, которым Abs могут подвергаться в процессе своего внутриклеточного созревания, N-связанное гликозилирование является одним из наиболее охарактеризованных.Широко признано, что Ab, экспрессируемые в клетках CHO, дифференциально гликозилированы по сравнению с Ab, продуцируемыми в других клетках млекопитающих (Routier et al. 1997; Sheeley et al. 1997). Миранда и др. (Miranda et al. 2007) показали, что Ab, продуцируемые клетками CHO, гликозилированы в большей степени, чем родительские Ab, продуцируемые гибридомой. Было обнаружено, что Ab F240, продуцированное в клетке CHO, имеет значительно больше концевых гликанов и сердцевинной фукозы по сравнению с Ab F240 из гибридомы.Более того, обработка пептидом N-гликозидазой F отменяла нейтрализацию F240. Это исследование показало, что структурные изменения, вызванные дополнительным N-гликозилированием в F240 Ab, определяют функциональные изменения его нейтрализующей активности.
Два других примера показывают влияние O-гликозилирования на конформацию и биологическую активность двух других белков, кальцитонина и прионного белка. Используя различные O-гликозилированные производные кальцитонина, каждое из которых имеет один остаток GalNAc, присоединенный к Ser или Thr, Tagashira et al.(Tagashira et al. 2001) продемонстрировали, что конформация кальцитонина и его гипокальциемическая активность сильно зависят от O-гликозилирования. Более того, эти авторы доказали, что эффект гликозилирования явно сайт-специфичен: гликозилирование по Thr-6 влияет как на структуру, так и на активность, только на активность Ser-5, а на Ser2 нет, указывая тем самым, что зависимость от сайта была очень строгой. Резкое различие в гипокальциемической активности, наблюдаемое в этом исследовании, можно объяснить стерическим ингибированием взаимодействия с рецептором (Tagashira et al.2001).
Уже давно известно, что структурный переход от PrP-c, нормального продукта прионного гена, к PrP-sc, патологической форме белка, приводит к образованию прионов и, однако, к заболеванию. механизм этого перехода был затемнен. На основании исследований прионных пептидов Chen et al. (Chen et al. 2002) предоставили доказательства связи O-гликозилирования со структурным переходом PrP-c в PrP-sc. Они обнаружили, что O-связанный α-GalNAc в Ser-135 подавляет развитие образования амилоидных фибрилл прионного пептида при физиологической концентрации соли, тогда как пептид с тем же сахаром, но в другом месте, в Ser-132, показал противоположное. эффект.Более того, этот эффект был специфичным для сахара: замена α-GalNAc на β-GalNac не давала такого же эффекта (Chen et al. 2002).
Как мы уже отмечали, описано много типов посттрансляционных модификаций белков (Rehfeld and Goetze, 2003). Все они направлены на изменение структуры родительской полипептидной цепи, что увеличивает количество функций белка. Очевидно, что apriori для достижения максимальной функциональной изменчивости и адаптивности к окружающей среде с помощью посттрансляционной модификации, компактная структура полипептида должна содержать сайты, которые могут быть чувствительны к различным типам модификации.Более того, один и тот же сайт может быть чувствительным к разным модифицирующим реакциям, поэтому разные модифицирующие агенты могут конкурировать за эти сайты (конкурентная модификация). Правильность такого предположения была подтверждена открытием, что фосфорилирование и гликозилирование могут конкурировать и изменять одни и те же аминокислотные остатки в целевых белках (Kelly et al. 1993).
O-связанный β-N-ацетилглюкозамин (O-GlcNAc) представляет собой модификацию моносахарида, в большом количестве присутствующую на остатках серина и треонина множества нуклео-цитоплазматических белков практически у всех высших эукариот, включая растения и грибы (Comer and Hart, 2000). ; Hart, 1997; Hart et al.2007; Торрес и Харт, 1984; Уэллс и Харт, 2003). Присоединение и удаление O-GlcNAc с помощью O-GlcNAc-трансферазы и O-GlcNAcase, соответственно, представляет собой быстрые реакции, аналогичные динамике добавления или удаления O-фосфата, контролируемой киназами и фосфатазами (Dong and Hart, 1994; Haltiwanger et al. 1992; Халтивангер и др., 1990; Кребс, 1993; Креппель и др., 1997; Креппель, Харт, 1999; Любас и др., 1997). Было показано, что сайты O-GlcNAc напоминают сайты фосфорилирования, и во многих случаях эти две модификации являются взаимоисключающими (Kelly et al.1993). Обратимое фосфорилирование и альтернативное гликозилирование вызывают изменения вторичных и третичных структур и контролируют функциональное поведение и модульные взаимодействия белков (Ahmad et al. 2007; Cheng and Hart, 2001; Pawson and Gish, 1992). Например, в случае рецептора эстрогена β (Cheng et al. 2000), большого Т-антигена SV-40 (Medina et al. 1998) и онкогена c-Myc (Chou et al. 1995a; Chou et al. 1995b), O-GlcNAc и O-фосфат конкурируют за один и тот же гидроксильный фрагмент. Как и фосфорилирование, O-GlcNAcylation реагирует на клеточный цикл, внеклеточные сигналы, метаболизм глюкозы, рост клеток и регуляцию транскрипции и трансляции (Chou and Omary, 1994; Datta et al.1989; Du et al. 2000; Han et al. 2000; Кирс и Харт, 1991; Roos et al. 1997; Roquemore et al. 1996).
Примеры, описанные выше, еще раз показывают, что первичный белок имеет много возможностей для модификации. Какой тип модификации он будет претерпевать, зависит от многих факторов, таких как тип клетки, клеточный цикл, внеклеточные и внутриклеточные сигналы, градиент pH, локальные концентрации конкретных модифицирующих молекул и т. Д. В результате белок будет выполнять определенную функцию в соответствующей клетке. , в определенном отсеке клетки и в точное время.
3. Пальмитоилирование белка
Следующий тип модификации белка, который мы хотели бы обсудить, — это S-пальмитоилирование. Основным механизмом, с помощью которого клетки регулируют субклеточную локализацию белков, является посттрансляционное липидирование (Draper et al. 2007). S-пальмитоилирование представляет собой одну из модификаций липидов, которая представляет собой обратимое тиоэфирное связывание пальмитата, 16-углеродной насыщенной жирной кислоты, с остатками цистеина (S-пальмитоилирование), катализируемое мембраносвязанными пальмитоил-трансферазами (PAT).Пальмитоилирование увеличивает гидрофобность модифицируемых молекул (Greaves and Chamberlain, 2007).
Это apriori очевидно, что одновременное проявление принципов структурной компактности с максимальной функциональной вариабельностью требует, чтобы сайты в молекуле полипептида, которые могут подвергаться различным типам модификации, перекрывались или даже были одинаковыми, и чтобы разные модифицирующие агенты были конкурируют за эти участки белков, изменяя их конформацию.Изменения в структуре белков способствует их локализации в определенных клеточных компартментах, способствует их мембранной ассоциации и контролирует межбелковые взаимодействия. Они также регулируют субклеточный перенос белков между мембранными органеллами и внутри микродоменов одних и тех же мембранных компартментов и модулируют их функции (Bijlmakers and Marsh, 2003; Casey and Seabra, 1996; Dunphy and Linder, 1998; el-Husseini Ael and Bredt, 2002; Миллиган и др., 1995; Надольски, Линдер, 2007; Реш, 1999; Сильвиус, 2002).
Для многих пальмитоилированных белков период полужизни пальмитатных фрагментов значительно короче, чем у белков-субстратов, что указывает на то, что комплекс проходит циклы депальмитоилирования и репальмитоилирования. Обратимость пальмитоилирования предполагает, что это регулируемая модификация, во многом похожая на фосфорилирование белка. Депальмитоилирование и репальмитоилирование могут обеспечивать механизм для регулирования таких процессов, как связывание цитозольных белков с мембранами, сегрегация белков в микродомены и опосредование межбелкового взаимодействия.Для белков, участвующих в передаче сигнала, эти циклы могут быть вызваны активацией и контролем доступа к конкретным субстратам (Bijlmakers and Marsh, 2003; Casey and Seabra, 1996; Dunphy and Linder, 1998; el-Husseini Ael and Bredt, 2002; Milligan. и др., 1995; Надольски, Линдер, 2007; Реш, 1999; Сильвиус, 2002). Цикл пальмитата на каркасном белке PSD-95 может проиллюстрировать этот механизм. Пальмитоилирование позволяет PSD-95 группироваться в постсинаптической мембране. Блокирование пальмитоилирования PSD-95 приводит к потере рецепторов AMPA из этих доменов за счет быстрого эндоцитоза рецепторов AMPA, что требует депальмитоилирования PSD-95 (el-Husseini Ael and Bredt, 2002; Nadolski and Linder, 2007).
Несколько других примеров могут дополнительно проиллюстрировать роль пальмитоилирования в функционировании белка. Было показано, что пальмитоилирование контролирует регулятор передачи сигналов G-белка (RGS). Пальмитоилирование остатка Cys в RGS-боксе важно для активности RGS16 GAP и их способности регулировать G-сопряженную передачу сигналов в клетках млекопитающих. Ферментативное пальмитоилирование RGS16 приводит к внутреннему пальмитоилированию остатка Cys-98. Мутация этого остатка на аланин снижает активность GAP слитого белка 5-HT-1A / Gα (Osterhout et al.2003 г.).
S-пальмитоилирование, как показано в исследовании Marino et al. (Marino et al. 2006), могут модулировать локализацию и функции рецептора эстрогена α (ER α). В клетках, экспрессирующих ERα дикого типа, основная популяция рецепторов эстрогена была мембранно-ассоциированной, тогда как в клетках, экспрессирующих мутант ERα Cys447-> Ala, мембраносвязанный рецептор обнаружен не был. Более того, мутация ERα Cys447-> Ala не позволяет E2-индуцированной пролиферативной передаче сигналов через пути ERK / MAPK и P13K / AKT в раковых клетках человека (Acconcia et al.2005). В целом, S-пальмитоилирование делает возможной экстраядерную локализацию ERα и способствует передаче сигналов для E2-опосредованной пролиферации (Marino et al. 2006).
S-пальмитоилирование, подобное фосфорилированию и гликозилированию, является важной посттрансляционной модификацией, которая обеспечивает белки многими важными функциями и контролирует клеточные процессы, зависящие от этих функций. Таким образом, как и другие посттрансляционные модификации, пальмитоилирование выполняет основной принцип структурно-функциональных отношений белков: получение максимальной функциональной активности путем внесения минимальных, но специфических изменений в структуру белка.
4. Сворачивание белка
Архитектура белка открывает много способов для получения белками множества функций. Эволюция приводит множество примеров, подтверждающих это утверждение. В некоторых случаях эволюция может объединять белки, изменяя третичную структуру на четвертичную. Например, пять отдельных ферментов в бактерии E.coli , которые катализируют последовательные этапы пути биосинтеза ароматических аминокислот, соответствуют пяти участкам одного белка у гриба Aspergillius nidulans (Lesk, 2001).
Чтобы стать функционально активным, вновь синтезированная полипептидная цепь должна складываться в уникальную трехмерную структуру. Хотя эксперименты in vitro твердо установили, что информация о трехмерной структуре белка генетически детерминирована и содержится в его аминокислотной последовательности (Anfinsen, 1973; Dobson and Karplus, 1999; Nagradova, 2004), сворачивание белка в нельзя считать, что клетка возникла спонтанно (Наградова, 2004). Стало ясно, что многим белкам требуется помощь для укладки в клетке, и что это обеспечивается белками-помощниками (молекулярными шаперонами) (Bukau and Horwich, 1998; Coyle et al.1997; Fewell et al. 2001; Gutsche et al. 1999; Хартл и Хайер-Хартл, 2002).
Действие шаперонов основано на двух различных механизмах. Первый механизм состоит в поддержании полипептидной цепи в состоянии, способном к продуктивному фолдингу, который происходит самопроизвольно после высвобождения развернутых цепей в раствор. Таким образом, взаимодействие с шапероном не влияет на сворачивание per se (Наградова, 2004).
Второй механизм используется шаперонами, чьи большие цилиндрические комплексы создают физически изолированные компартменты, предназначенные для сворачивания полипептидов, частично свернутых или неправильно свернутых белков, которые инкапсулируются внутри центральной полости.Поскольку внутри полости может поместиться только одна молекула белка-субстрата, ее складывание происходит в условиях, полностью исключающих агрегацию (Наградова, 2004). Однако роль шаперонов явно не ограничивается их способностью создавать изолированные компартменты, где белок может спонтанно складываться в условиях, имитирующих бесконечное разведение (модель «клетки Анфинсена») (Ellis and Hartl, 1996). На сегодняшний день накоплен большой объем информации, подтверждающий представление о том, что шапероны могут играть активную роль в сворачивании белков, ускоряя этот процесс и значительно повышая его эффективность (Наградова, 2004).Специальная группа белков-помощников, так называемых фолдаз (Balbach, 2000; Wu and Matthews, 2002) и дисульфид-изомераз (Creighton, 2002; Frand et al. 2000), способствует укладке белков, катализируя лимитирующие скорость реакции изомеризации.
5. Сплайсинг белков
Простое представление об экспрессии белка постоянно подвергается сомнению новыми примерами посттрансляционного процессинга. Эти примеры включают нерибосомное добавление фрагментов и протеолитическое расщепление полипротеина, а также вырезание и перестройки, такие как опосредованный интеином сплайсинг белка, сплайсинг путем обратного протеолиза, авто-процессинг белка и опосредованное протеасомами лигирование пептидов (David et al.2004; Hanada et al. 2004; Норен и др. 2000; Паулюс, 2001; Перлер, 2005; Perler et al. 1994; Vigneron et al. 2004 г.).
a) Интеин-опосредованный сплайсинг белков
Интеины — это внутренние сегменты белков-предшественников, которые катализируют их удаление ipso с помощью внутримолекулярного процесса, называемого сплайсингом белков, с одновременным лигированием двух фланкирующих внешних областей (N- и C-экстеинов. ) через нативную пептидную связь (David et al. 2004; Perler, 2005). Интеины обнаружены в белках одноклеточных организмов, принадлежащих ко всем трем доменам жизни, архей, бактерий и эукариот, а также в вирусных и фаговых белках, но не были обнаружены у высших организмов, таких как позвоночные (Hanada and Yang, 2005).Известно более 100 интеинов. Белки-хозяева интеина очень разнообразны, включая полимеразы ДНК и РНК, рибонуклеотидредуктазы, АТФазы, протеазы, метаболические ферменты, факторы транскрипции и трансляции (Pietrokovski, 2001). Следующие свойства отличают опосредованный интеином белковый сплайсинг от других типов белковых перестроек: (а) сигнатурная последовательность интеина, (b) самокаталитическая реакция в отсутствие экзогенных белков, кофакторов или источников энергии, и (c) образование нативная пептидная связь между лигированными экстеинами (Perler, 2005).
Интеины можно разделить на четыре класса: макси-интеины (со встроенным эндонуклеазным доменом), мини-интеины (без эндонуклеазного домена), интеины транс-сплайсинга (где сплайсинговые соединения не связаны ковалентно) и интеины Ala (Ala как N-концевая аминокислота) (Evans, Xu, 2002). Последовательности, специфичные для интеина, имеют некоторые общие характеристики. Они появляются в консервативных областях белка-хозяина. Все последовательности интеина несут разные блоки, обозначенные A и B в N-концевом домене сплайсинга и F и G в C-концевом домене сплайсинга.
Интеины обычно имеют N-концевые остатки Cys, Ser или Ala и C-концевые остатки His или Asn и вставлены в белки-хозяева рядом с Cys, Ser или Tre (Hanada et al. 2004). Хотя указанные выше аминокислоты являются типичными для интеинов, многие интеины имеют другие аминокислоты в определенных блоках. Вероятно, что каждый интеин имеет определенную комбинацию нуклеофилов и вспомогательных групп, отточенных во время эволюции для оптимальной функции. Структурные исследования не выявили большего количества остатков, способствующих консенсусу, потому что каждый интеин имеет разные остатки со сходными химическими функциями в активном сайте или рядом с ним (Perler, 2005).Наименьший известный интеин состоит из 134 аминокислот (Evans et al. 1999), а большинство из них намного больше.
Роль интеинов в клетке пока неясна, однако можно предположить, что, поскольку интеины интегрированы в высококонсервативные области белка, функция (и) белка-хозяина до и после удаления интеина может быть различной. Если интеин должен вырезать себя для активации функции белка-хозяина, он должен быть чувствительным к специфическим сигналам. Есть некоторые признаки того, что это так (Chong et al.1998; Саутворт и др. 1999). Установлено, что интеин регулирует активность ДНКЕ Synechocystis sp.PCC6803, функционирующей как внутренний регулятор (Ghosh et al. 2001). Свободные интеины (после сплайсинга) могут выполнять как регуляторные, так и ферментативные функции сами по себе (Klabunde et al. 1998; Starokadom-skyy, 2007). Таким образом, хотя это было доказано только для ограниченного числа белков, интеины могут определять различные функции белка-хозяина до и после сплайсинга и получать новую функцию (и) после самоэкцизии.
b) Посттрансляционный сплайсинг посредством обратного протеолиза
Другой пример посттрансляционной модификации, сплайсинга белков посредством обратного протеолиза, представляет собой сплайсинг белков, обнаруженный у растений. Это наблюдалось с лектином конканавалином A (Con A) (Carrington et al. 1985). Были описаны следующие этапы такого сплайсинга белков. Первоначальный предшественник Con A (гликопро-Con A) сначала активируется дегликозилированием до про-Con A. Затем Pro-Con A расщепляется с образованием двух различных белков, которые транспонируются и превращаются в зрелый Con A.Хотя точный механизм этого сплайсинга у растений все еще не ясен, исследование in vitro показало, что аспарагинилэндопептидаза может переваривать Con A, а затем повторно лигировать расщепленные фрагменты за счет своей обратной протеолитической активности (Min and Jones, 1994).
c) Сплайсинг белков, опосредованный протеасомами
Недавно было показано, что протеасомы представляют собой клеточные компартменты, в которых происходит не только деградация белка, но и посттрансляционные модификации белков (Hanada and Yang, 2005; Hanada et al.2004; Vigneron et al. 2004 г.). Hanada et al. (Hanada et al. 2004) обнаружили, что созревание и презентация антигена могут осуществляться в протеасомах путем сплайсинга белков, включающего вырезание пептида и повторное лигирование. Они показали, что в отличие от сплайсинга интеинов, когда обычно вырезаемый полипептид имеет длину не менее 134 аминокислот (наименьший известный интеин состоит из 134 аминокислот), вырезанные полипептиды в их системе имели длину всего 18-40 аминокислот. Используя ингибитор протеасом, clasto-lactacystin β-lactone, авторы доказали, что сплайсинг белка FGF-5, наблюдаемый в их системе, был опосредован протеасомами (Hanada and Yang, 2005).Более того, эти авторы впервые показали, что сплайсинг белков может происходить в клетках человека.
Другая группа также описала продукцию антигена из белка путем сплайсинга несмежных пептидов в обратном порядке (Vigneron et al. 2004). Этот сплайсинг был проведен 20S протеасомой in vitro и in vivo . В описанных системах длина вырезанной промежуточной области составляла всего четыре аминокислоты (Noren et al. 2000; Vigneron et al. 2004). Таким образом, опосредованный протеасомами сплайсинг преодолевает ограничение, связанное с длиной промежуточных последовательностей, которое имеет решающее значение для опосредованной интеином модификации белка.
Каковы последствия этих выводов? Как сообщает Hanada et al. подчеркнули, что эти результаты показывают, что «иммунная система отслеживает несмежные пептидные последовательности, генерируемые посттрансляционно. Эта способность представляет собой огромное увеличение способности CTL распознавать собственные и внешние белки… В более широком контексте существование сплайсинга белков у позвоночных значительно увеличивает возможности клетки для преобразования генетической информации в белки »(Hanada et al. 2004). Эти результаты также показывают, что сплайсинг белков является мощным механизмом, позволяющим реализовать главный принцип многофункциональности белков — преобразование компактной генетической информации в широкое разнообразие структур и функций белка.«Уже было известно, что такие процессы, как рекомбинация ДНК и сплайсинг РНК, увеличивают количество различных белков, продуцируемых каждым из генов млекопитающих; открытие сплайсинга белков дополняет набор инструментов »(Rammensee, 2004).
Функции белков | 5 основных функций, что, где и как
Белки — это самые распространенные органические молекулы, присутствующие на Земле. Они в изобилии присутствуют в каждой живой клетке. Белки — это полимеры, состоящие из тысяч аминокислот, связанных пептидными связями.Длинные цепи аминокислот, известные как полипептиды, складываются вокруг себя несколькими способами, образуя сложные структуры, называемые белками.
Функции, выполняемые белками, можно разделить на разные категории. Некоторые функции важны на клеточном уровне, в то время как другие необходимы для лучшей работы организма в целом. Здесь мы попытаемся понять различные функции, выполняемые белками в нашем организме, на различных примерах.
Все ферменты — это белки
Ферменты — это белки, которые необходимы для любой химической реакции в нашем организме.Они катализируют биохимическую реакцию, чтобы жизнь продолжалась.
Пример ферментативной реакции в нашем организме — гликолиз. Это процесс высвобождения энергии из молекулы глюкозы. Эта энергия требуется для выполнения нескольких процессов, происходящих внутри клетки. Процесс гликолиза включает около 10 этапов, каждый из которых требует определенного фермента. Отсутствие единственного фермента останавливает процесс, и энергия из глюкозы не может быть получена.
Для синтеза белков также требуются определенные ферменты.Синтез белка включает транскрипцию ДНК в мРНК, а затем трансляцию мРНК рибосомами. Оба эти шага требуют ферментов, которые являются белками. Например;
РНК-полимераза — это фермент, необходимый для соединения нуклеотидов РНК в процессе транскрипции.
Аминоацил тРНК синтетаза — это фермент, который присоединяет определенные аминокислоты к тРНК, чтобы ее можно было использовать в синтезе белка.
Таким образом, от получения энергии до производства белков, все химические процессы в живых организмах нуждаются в ферментах, и все ферменты являются белками.Роль белков как ферментов — самая важная и важная функция, выполняемая белками.
Белки действуют как рецепторы на клеточных мембранах
Белки являются важными компонентами всех клеточных мембран и мембран органелл. Одна из функций этих мембранных белков состоит в том, что они действуют как рецепторы. Гормоны, нейротрансмиттеры и другие сигнальные молекулы связываются с этими рецепторами и передают сигналы клеткам. Таким образом, белки играют роль в передаче сигналов клетками, которая важна для скоординированной функции всех клеток, присутствующих в нашем организме.Рассмотрим следующий пример, чтобы понять роль белков как рецепторов.
Инсулин — это гормон, контролирующий уровень глюкозы в крови. Он выполняет свою функцию, связываясь со своим рецептором, который является белком. Инсулин связывается со своим рецептором, который посылает сигналы для открытия каналов глюкозы, чтобы глюкоза могла поступать из крови в печень и мышечные клетки. Если рецепторы инсулина отсутствуют, уровень глюкозы в крови нельзя регулировать.
Этот и ряд других примеров в нашем организме доказывают, почему белки необходимы для передачи сигналов клетками и координации клеточных функций.
Некоторые гормоны также являются белками
Белки действуют не только как клеточные рецепторы, но и как гормоны. Инсулин и глюкагон — два гормона, которые по своей природе являются белками. Оба эти гормона необходимы для регулирования уровня глюкозы в крови. Они контролируют поглощение и высвобождение глюкозы клетками, гликолиз и глюконеогенез, а также синтез и деградацию гликогена. Роли этих гормонов в нашем организме перечислены ниже;
Инсулин вырабатывается поджелудочной железой при высоком уровне глюкозы в крови.Он способствует усвоению клетками глюкозы, ее расщеплению, а также хранению в виде гликогена. Он также подавляет синтез новых молекул глюкозы из неуглеводных источников (глюконеогенез).
Глюкагон высвобождается поджелудочной железой при низком уровне глюкозы в крови. Он способствует расщеплению гликогена с высвобождением глюкозы. Он также способствует глюконеогенезу.
Белки действуют как транспортные каналы в клеточных мембранах
Белки, присутствующие в клеточных мембранах, также действуют как транспортные каналы.Вещества, которые не проницаемы через мембраны из-за своего размера или заряда, могут проникать в клетку через эти белковые каналы. Один белковый канал специфичен для одного или нескольких веществ. Примеры белковых каналов приведены ниже;
Аквапорины — это белковые каналы, которые позволяют молекулам воды проходить через клетки
Натриевые каналы позволяют проходить ионам натрия внутри клетки
Калиевые каналы пропускают только калий ионы проходят через них
Кальциевые каналы специфичны только для ионов кальция
Это несколько примеров белковых каналов, присутствующих в мембранах.
Белки поддерживают форму и структуру клетки
Это еще одна важная клеточная функция, выполняемая белками. Цитоскелет состоит из нескольких связанных между собой белковых нитей. Белки в цитоскелете организованы в виде микротрубочек, микрофиламентов и промежуточных филаментов. Все эти компоненты цитоскелета расположены определенным образом, сохраняя форму клетки. Важные белки, из которых состоит цитоскелет, включают актин и тубулин.В отсутствие этих белков клетка не могла бы поддерживать свою структуру.
Белки участвуют в делении клеток
Деление клеток — это процесс, посредством которого зрелая взрослая родительская клетка делится на дочерние клетки. Белки также необходимы для этого процесса.
Во время деления клетки хромосомы разделяются на две половины путем растяжения. Это разделение хромосом осуществляется белками, известными как волокна веретена.
Белки также необходимы для деления цитоплазмы, которое происходит после разделения хромосом.
Белки необходимы для транспорта внутри клетки
Определенные транспортные белки необходимы для внутриклеточного транспорта различных веществ. Различные белки, которые участвуют во внутриклеточных белках, известны как моторные белки. Эти белки используют энергию в форме АТФ и перемещаются по микротрубочкам для транспортировки различных веществ в цитоплазме клетки. Примером моторных белков является белок кинезин. Он участвует в транспорте различных веществ в аксонах нейронов.
Белки необходимы для транспорта кислорода
Эта функция белков важна для выживания организма в целом. В этом процессе участвуют два белка — гемоглобин и миоглобин.
Гемоглобин
Это белок, присутствующий в красных кровяных тельцах. Гемоглобин состоит из четырех полипептидных цепей, двух альфа-цепей и двух бета-цепей, которые намотаны друг на друга. Каждая из этих полипептидных цепей несет одну гемовую группу (содержащую атом железа).
Этот белок отвечает за перенос кислорода из легких в тканевую жидкость. Одна молекула кислорода может связываться с четырьмя молекулами кислорода. Он связывается с молекулами кислорода, присутствующими в воздухе, проходя через легкие. Эти молекулы кислорода высвобождаются, когда кровь проходит через ткани.
Любой недостаток или отклонение от нормы гемоглобина нарушает перенос кислорода кровью. Наши клетки не могут выжить без кислорода. Любое нарушение подачи кислорода приведет к гибели клеток в пораженных тканях.
Миоглобин
Миоглобин — еще один белок, участвующий в транспортировке кислорода. Он состоит из одной полипептидной цепи с гемовой группой. Это цитоплазматический белок, имеющий более высокое сродство к молекулам кислорода, что означает, что он может связываться с кислородом даже при высокой концентрации кислорода. Его функция — переносить кислород из тканевой жидкости в клетки.
Из-за своего высокого сродства к кислороду миоглобин выделяет кислород в очень низких концентрациях.Эта особенность миоглобина отвечает за хранение кислорода в тканях.
Белки необходимы для транспортировки различных веществ в крови
Хотя кровь действует как транспортная среда, белки необходимы для удержания и транспортировки некоторых веществ, которые не могут растворяться в крови. Эта функция белков также важна для правильного функционирования организма. Некоторые примеры транспортных белков, присутствующих в крови, следующие.
Альбумин является основным транспортным белком крови.Он действует как переносчик жирных кислот, стероидов, гормонов щитовидной железы, липофильных препаратов, тяжелых металлов, ионов кальция и билирубина
Преальбумин — еще один транспортный белок в крови, который переносит стероидные гормоны, тироксин и витамин A
Гаптоглобин является транспортным средством белок, несущий свободный гемоглобин, присутствующий в плазме
Связывающий тироксин белок специфичен для гормона щитовидной железы
ЛПВП — это липопротеин, который транспортирует холестерин из тканей в печень
ЛПНП — еще один липопротеин, который транспортирует холестерин из печени в ткани
Белки участвуют в сокращении мышц
Сокращение мышц — это процесс, который позволяет нам выполнять повседневные жизненные задачи, такие как ходьба, бег, сидение, стояние, письмо и даже речь.Этот процесс сокращения мышц также происходит из-за белков. Сократительные белки присутствуют в мышечных волокнах. Эти белки взаимодействуют особым образом, что позволяет сокращать и расслаблять мышцы. Самыми важными сократительными присутствующими являются:
Белки предотвращают отек
Отек — это состояние, при котором избыточная жидкость вытекает из кровеносных сосудов и собирается в тканевых пространствах. Потеря жидкости из крови приводит к снижению артериального давления. Это потенциально смертельное состояние, которое может поставить под угрозу эффективную доставку крови к тканям организма.
Белки, присутствующие в крови, известные как белки плазмы, предотвращают утечку жидкости через капилляры благодаря своему осмотическому эффекту. Онкотическое давление из-за белков плазмы удерживает воду внутри кровеносных сосудов, предотвращая ее попадание в тканевые жидкости, предотвращая отек. Если эти белки отсутствуют, отек развивается в разных частях тела.
Белки защищают наш организм от болезней
Эту функцию выполняют антитела. Антитела или иммуноглобулины — это белки плазмы, которые вырабатываются в ответ на попадание в наш организм различных болезнетворных агентов.Они борются с этими патогенами и защищают наш организм от их вредного воздействия. Если в нашем организме уже присутствуют антитела против патогена, они уничтожают патоген до того, как он вызовет какое-либо заболевание. Этот процесс известен как иммунитет.
Белки необходимы для пищеварения
Процесс пищеварения включает расщепление сложных веществ, присутствующих в нашем рационе, на более простые, чтобы они могли всасываться в кровь. Расщепление различных пищевых веществ на более простые молекулы происходит в нашей пищеварительной системе ферментами, которые по своей природе являются белками.
Белки также действуют как запасные вещества
Белки представляют собой полимеры аминокислот. Они действуют как запасные вещества, в которых хранятся тысячи аминокислот. Эти аминокислоты высвобождаются из белков, когда они необходимы организму. Примеры запасных белков:
Казеин в молоке
Альбумин в яйце
Эти белки обеспечивают незаменимые аминокислоты, необходимые организму для производства нескольких белков. Более того, во время голодания белки, присутствующие в организме, также могут использоваться в качестве источника энергии для обеспечения калорий, необходимых для выполнения различных функций организма.
Белки контролируют экспрессию генов
Экспрессия генов — это процесс, посредством которого информация в конкретном гене копируется в форме мРНК, а позже эта мРНК используется рибосомами для создания белка, кодируемого этим геном.
Этот процесс экспрессии генов контролируется факторами транскрипции. Эти факторы транскрипции позволяют транскрипцию генов только тех белков, которые в настоящее время необходимы организму.
Факторы транскрипции также являются белками по своей природе.Таким образом, белки регулируют свой собственный синтез, регулируя экспрессию генов.
Резюме
Белки — это полимеры, состоящие из аминокислот. Они участвуют практически во всех процессах, происходящих в нашем организме. Сводка функций, выполняемых белками, выглядит следующим образом;
Как ферменты, белки необходимы для всех химических процессов в живых организмах
Как гормоны и клеточные рецепторы, они необходимы для клеточной передачи сигналов и координации
Как транспортные каналы, белки необходимы для проникновения ионов и более -размерные частицы в клетки
Являясь компонентами цитоскелета, они поддерживают форму клеток
Волокна веретена — это белковые волокна, которые необходимы для деления клеток
Гемоглобин и миоглобин являются белками, необходимыми для транспорта кислорода
Альбумин и другая плазма белки необходимы для транспорта липидов, лекарств и других веществ в крови
Сократительные белки необходимы для сокращения мышц
Антитела — это белки, которые защищают наш организм от вредоносных болезней
Белки плазмы поддерживают баланс жидкости в нашем организме
Они регулируют экспрессию генов
Белки также обеспечивают эргия тела во время голода
Ссылки
Lodish H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipurksy SL, Darnell J (2004).Молекулярная клеточная биология (5-е изд.). Нью-Йорк, Нью-Йорк: WH Freeman and Company
Zhang C, Kim SH (февраль 2003 г.). «Обзор структурной геномики: от структуры к функции» . Текущее мнение в химической биологии. 7 (1): 28–32. doi : 10.1016 / S1367-5931 (02) 00015-7 . PMID 12547423
Sleator RD (2012).«Прогнозирование функций белков». Функциональная геномика. Методы молекулярной биологии. 815 . С. 15–24. doi : 10.1007 / 978-1-61779-424-7_2 . ISBN 978-1-61779-423-0 . PMID 22130980
Роль мембранных белков в физиологии
Мембранные белки — это белки, которые являются частью клеточных мембран или взаимодействуют с ними, и они несут ответственность за выполнение большинства функций этих мембран.Мембранные белки составляют примерно одну треть белков человека и отвечают за регулирование процессов, которые помогают биологическим клеткам выжить.
Изображение предоставлено: sciencepics / Shutterstock.com
Мембранные белки имеют ряд различных структур и также расположены в разных областях мембраны. Они выполняют широкий спектр функций, и количество белков и типы белков, присутствующих на конкретной мембране, могут варьироваться.
Структура мембранного белка
Клеточные мембраны состоят из двух фосфолипидных бислоев, которые называются листочками.Эти листочки присутствуют на всех клетках, образуя барьер, окружающий каждую клетку. Мембранные белки находятся на этих бислоев фосфолипидов или они взаимодействуют с этими бислоями фосфолипидов.
Внутри липидного бислоя находятся неполярные мембранные белки, которые являются гидрофобными (водоотталкивающими), и полярные мембранные белки, которые являются гидрофильными (способны смешиваться с водой). Они напрямую связаны с липидными бислоями, которые создают барьер вокруг каждой клетки.
Интегральные мембранные белки прочно закреплены на мембране.
Белки периферической мембраны не являются постоянной частью мембраны и могут иметь гидрофобные, электростатические и другие нековалентные взаимодействия с мембраной или интегральными белками.
Интегральные белки бывают разных типов, таких как монотопные, битопические, политопные, липидно-заякоренные белки или трансмембранные белки.
Монотопные интегральные белки прикреплены только к одному из двух листочков клетки.
Интегральные белки Bitopic — это трансмембранные белки, которые могут однажды покрывать липидные бислои.Политопные белки также являются трансмембранными белками, которые охватывают липидные бислои более одного раза.
Заякоренный липидом белок имеет ковалентную связь с липидами, которые встроены в фосфолипидный бислой.
Функции мембранного белка
Существует множество функций, которые выполняют мембранные белки. К ним относятся:
Соединения: соединение двух ячеек вместе
Ферментативные функции
Все ферменты представляют собой белок.В результате мембранный белок, внедренный в мембрану, иногда может быть ферментом, активный сайт которого может быть обращен к веществам за пределами липидного бислоя.
Эти типы ферментативных мембранных белков могут работать в группах для выполнения определенных этапов метаболизма, например, расщепления лактозы на углеводы, а затем на моносахариды.
Транспорт
Мембранные белки могут позволить гидрофильным молекулам проходить через клеточную мембрану.Белки транспортной мембраны бывают разных форм, и некоторым требуется энергия для изменения формы и активного перемещения молекул и других веществ через клеточную мембрану. Они делают это, высвобождая АТФ для использования в качестве источника энергии.
Крепление: становятся точками прикрепления цитоскелета и внеклеточного матрикса
Преобразование сигнала
Некоторые мембранные белки могут иметь сайт связывания. Эти сайты связывания имеют особую форму, которая соответствует форме химического посредника.Например, эти химические посланники могут быть гормонами.
Когда гормон встречается с клеточной стенкой, он соединяется с рецепторным мембранным белком, встроенным в клеточную стенку. Гормон может изменить рецепторный белок и вызвать определенную реакцию, в зависимости от типа гормона или другого вещества, которая будет иметь место внутри клетки.
Распознавание клеток
Другая важная функция мембранных белков — идентификация и распознавание между клетками.Эта конкретная функция полезна для иммунной системы, поскольку она помогает организму распознавать чужеродные клетки, например, которые могут вызывать инфекцию. Гликопротеины — это один из типов мембранных белков, которые могут распознавать клетки.
Межклеточное соединение
Соседние клетки могут иметь мембранные белки, которые соединяются в различных соединениях. Щелевые и плотные соединения.
Эта функция помогает клеткам общаться друг с другом и передавать материалы между собой.
Приложение
Мембранные белки важны для цитоскелета, системы нитей и волокон в цитоплазме клетки и внеклеточного матрикса (ECM), который представляет собой сеть макромолекул, находящихся вне клеток, таких как коллаген, ферменты и гликопротеины. к мембранным белкам.
Прикрепление нитей или волокон в цитоплазме по всей клетке может помочь клетке сохранять свою особую форму. Он также сохраняет стабильное расположение мембранных белков.
Присоединение мембранных белков к внеклеточному матриксу может помочь ЕСМ опосредовать изменения, происходящие во внеклеточной и внутриклеточной среде.
Мембранные белки при болезнях
Некоторые заболевания связаны с мутациями мембранных белков. Одним из примеров является мутация под названием V509A, обнаруженная в рецепторе тиреотропина, причем тиреотропин является гормоном, секретируемым гипофизом, который регулирует выработку гормонов щитовидной железы.
Эта мутация увеличивает активность рецептора тиреотропина и приводит к врожденному гипертиреозу, состоянию, которое может вызывать изменения настроения, проблемы со сном и проблемы с желудком.
Другие заболевания, связанные с мутациями в мембранных белках, включают наследственную глухоту, болезнь Шарко-Мари-Тута, которая повреждает периферические нервы вне центральной нервной системы, и синдром Дежерина-Соттаса, который влияет на способность человека двигаться.
Изображение предоставлено: Explode / Shutterstock.com
Сводка
Мембранные белки выполняют ряд важных функций, которые помогают клеткам общаться, сохранять свою форму, выполнять изменения, вызванные химическими посредниками, а также транспортировать и обмениваться материалом.
Мембранные белки также могут играть роль в прогрессировании заболевания, поскольку иммунная система может использовать мембранные белки для идентификации потенциально вредных чужеродных молекул в организме.
Функции белков в организме — Питание человека [УСТАРЕЛО]
Белки являются «рабочими лошадками» тела и участвуют во многих функциях организма. Белки бывают всех размеров и форм, и каждый из них специально структурирован для своей конкретной функции.
Структура и движение
Рисунок 6.9 Структура коллагена
Тройная спираль коллагена от Невита Дилмена / CC BY-SA 3.0
В организме человека было обнаружено более сотни различных структурных белков, но наиболее распространенным является коллаген, который составляет около 6 процентов от общей массы тела. Коллаген составляет 30 процентов костной ткани и включает большое количество сухожилий, связок, хрящей, кожи и мышц. Коллаген — это прочный волокнистый белок, состоящий в основном из глицина и пролина. Внутри его четвертичной структуры три пептидных нити скручиваются друг с другом, как веревка, а затем эти коллагеновые нити перекрываются друг с другом.Эта высокоупорядоченная структура даже прочнее, чем стальные волокна того же размера. Коллаген делает кости крепкими, но гибкими. Волокна коллагена в дерме кожи придают ей структуру, а сопутствующие фибриллы белка эластина делают ее гибкой. Зажмите кожу на руке и отпустите; белки коллагена и эластина в коже позволяют ей вернуться к своей первоначальной форме. Гладкомышечные клетки, которые выделяют белки коллагена и эластина, окружают кровеносные сосуды, придавая им структуру и способность растягиваться назад после того, как через них прокачивается кровь.Еще один сильный волокнистый белок — кератин, из которого состоят кожа, волосы и ногти. Плотно упакованные коллагеновые фибриллы в сухожилиях и связках обеспечивают синхронные механические движения костей и мышц и способность этих тканей возвращаться назад после завершения движения.
Ферменты
Хотя белки в наибольшем количестве содержатся в соединительных тканях, таких как кости, их наиболее необычная функция — это ферменты. Ферменты — это белки, которые проводят определенные химические реакции.Задача фермента — обеспечить место для химической реакции и снизить количество энергии и время, необходимое для того, чтобы эта химическая реакция произошла (это известно как «катализ»). В среднем каждую секунду в клетках происходит более ста химических реакций, и для большинства из них требуются ферменты. Одна только печень содержит более тысячи ферментных систем. Ферменты специфичны и будут использовать только определенные субстраты, которые подходят их активному сайту, подобно тому, как замок может быть открыт только с помощью определенного ключа.Почти каждая химическая реакция требует определенного фермента. К счастью, фермент может снова и снова выполнять свою роль катализатора, хотя в конечном итоге он разрушается и восстанавливается. Все функции организма, включая расщепление питательных веществ в желудке и тонком кишечнике, преобразование питательных веществ в молекулы, которые клетка может использовать, и построение всех макромолекул, включая сам белок, включают ферменты (см. Рисунок 6.10 «Роль ферментов в переваривании углеводов») .
Рисунок 6.Роль 10 ферментов в переваривании углеводов
Гормоны
Белки отвечают за синтез гормонов. Гормоны — это химические сообщения, производимые железами внутренней секреции. Когда эндокринная железа стимулируется, она выделяет гормон. Затем гормон транспортируется с кровью к своей клетке-мишени, где он передает сообщение, чтобы инициировать определенную реакцию или клеточный процесс. Например, после еды уровень глюкозы в крови повышается. В ответ на повышение уровня глюкозы в крови поджелудочная железа выделяет гормон инсулин.Инсулин сообщает клеткам организма, что глюкоза доступна и может забирать ее из крови и хранить или использовать для производства энергии или создания макромолекул. Основная функция гормонов — включать и выключать ферменты, поэтому некоторые белки могут даже регулировать действие других белков. Хотя не все гормоны состоят из белков, многие из них таковы.
Жидкостный и кислотно-щелочной баланс
Правильное потребление белка позволяет основным биологическим процессам организма поддерживать статус-кво в изменяющейся окружающей среде.Баланс жидкости относится к поддержанию распределения воды в организме. Если слишком много воды в крови внезапно попадает в ткань, это приводит к отеку и, возможно, к гибели клеток. Вода всегда течет из области с высокой концентрацией в область с низкой концентрацией. В результате вода перемещается в области с более высокими концентрациями других растворенных веществ, таких как белки и глюкоза. Чтобы вода равномерно распределялась между кровью и клетками, белки постоянно циркулируют в крови в высоких концентрациях.Самый распространенный белок в крови — это белок в форме бабочки, известный как альбумин. Присутствие альбумина в крови делает концентрацию белка в крови похожей на таковую в клетках. Таким образом, обмен жидкости между кровью и клетками не является экстремальным, а скорее сводится к минимуму, чтобы сохранить статус-кво.
Рисунок 6.11 Белковый альбумин
PDB 1o9x EBI Джавахара Сваминатана и сотрудников MSD из Европейского института биоинформатики / Public Domain Белок в форме бабочки, альбумин, выполняет множество функций в организме, включая поддержание жидкостного и кислотно-щелочного баланса и транспортировку молекул.
Белок также необходим для поддержания правильного баланса pH (мера того, насколько кислым или основным является вещество) в крови. PH крови поддерживается между 7,35 и 7,45, что является слегка щелочным. Даже небольшое изменение pH крови может повлиять на функции организма. Напомним, что кислая среда может вызвать денатурацию белка, что останавливает функционирование белков. В организме есть несколько систем, которые удерживают pH крови в пределах нормы, чтобы этого не происходило. Один из них — это циркулирующий альбумин.Альбумин имеет слабую кислотность и, поскольку он отрицательно заряжен, уравновешивает множество положительно заряженных молекул, таких как протоны (H +), кальций, калий и магний, которые также циркулируют в крови. Альбумин действует как буфер против резких изменений концентраций этих молекул, тем самым уравновешивая pH крови и поддерживая статус-кво. Белок гемоглобин также участвует в кислотно-щелочном балансе, связывая и высвобождая протоны.
Транспорт
Альбумин и гемоглобин также играют роль в молекулярном транспорте.Альбумин химически связывается с гормонами, жирными кислотами, некоторыми витаминами, необходимыми минералами и лекарствами и переносит их по кровеносной системе. Каждый эритроцит содержит миллионы молекул гемоглобина, которые связывают кислород в легких и транспортируют его ко всем тканям организма. Плазматическая мембрана клетки обычно не проницаема для больших полярных молекул, поэтому для доставки необходимых питательных веществ и молекул в клетку в клеточной мембране существует множество транспортных белков. Некоторые из этих белков являются каналами, которые позволяют определенным молекулам входить и выходить из клеток.Другие действуют как такси с односторонним движением и требуют энергии для работы.
Защита
Рисунок 6.12 Белки антител
Абаговомаб (моноклональное антитело) по Blake C / CC BY-SA 3.0
Рисунок 6.13 Антигены
Цепи антител от Fred the Oyster / Public Domain
Белок антитела состоит из двух тяжелых цепей и двух легких цепей. Вариабельная область, которая отличается от одного антитела к другому, позволяет антителу распознавать соответствующий ему антиген.
Ранее мы обсуждали, что прочные волокна коллагена в коже обеспечивают ей структуру и поддержку.Плотная сеть коллагеновых волокон кожи также служит преградой для вредных веществ. Функции атаки и разрушения иммунной системы зависят от ферментов и антител, которые также являются белками. Фермент лизоцим выделяется со слюной и атакует стенки бактерий, вызывая их разрыв. Определенные белки, циркулирующие в крови, могут быть направлены на создание молекулярного ножа, который пронзает клеточные мембраны чужеродных захватчиков. Антитела, выделяемые лейкоцитами, исследуют всю систему кровообращения в поисках вредных бактерий и вирусов, которые можно окружить и уничтожить.Антитела также запускают другие факторы иммунной системы для поиска и уничтожения нежелательных злоумышленников.
Заживление ран и регенерация тканей
Белки участвуют во всех аспектах заживления ран, процесса, который проходит в трех фазах: воспалительной, пролиферативной и ремоделирующей. Например, если вы шили и укололи палец иглой, ваша плоть покраснела бы и воспалилась. Через несколько секунд кровотечение прекратится. Процесс заживления начинается с белков, таких как брадикинин, которые расширяют кровеносные сосуды в месте повреждения.Дополнительный белок, называемый фибрином, помогает защитить тромбоциты, которые образуют сгусток, чтобы остановить кровотечение. Затем, в фазе пролиферации, клетки перемещаются и восстанавливают поврежденную ткань, устанавливая новые коллагеновые волокна. Волокна коллагена помогают сблизить края раны. В фазе ремоделирования откладывается больше коллагена, образуя рубец. Рубцовая ткань только на 80 процентов функциональна, чем нормальная неповрежденная ткань. Если в диете недостаточно белка, процесс заживления ран заметно замедляется.
В то время как заживление ран происходит только после получения травмы, в организме продолжается другой процесс, называемый регенерацией тканей. Основное различие между заживлением ран и регенерацией тканей заключается в процессе восстановления точной структурной и функциональной копии утраченной ткани. Таким образом, старая умирающая ткань заменяется не рубцовой тканью, а совершенно новой, полностью функциональной тканью. Некоторые клетки (например, клетки кожи, волос, ногтей и кишечника) имеют очень высокую скорость регенерации, в то время как другие (например, клетки сердечной мышцы и нервные клетки) не регенерируют на каких-либо заметных уровнях.Регенерация тканей — это создание новых клеток (деление клеток), для чего требуется множество различных белков, включая ферменты, синтезирующие РНК и белки, транспортные белки, гормоны и коллаген. В волосяном фолликуле клетки делятся, и волосы растут в длину. Рост волос в среднем составляет 1 сантиметр в месяц, а ногтей — около 1 сантиметра каждые сто дней. Клетки, выстилающие кишечник, восстанавливаются каждые три-пять дней. Неадекватные белковые диеты ухудшают регенерацию тканей, вызывая множество проблем со здоровьем, включая нарушение переваривания и усвоения питательных веществ и, что наиболее заметно, роста волос и ногтей.
Производство энергии
Некоторые аминокислоты в белках можно разобрать и использовать для производства энергии (Рисунок 6.14 «Аминокислоты, используемые для получения энергии»). Только около 10 процентов пищевых белков катаболизируются каждый день для производства клеточной энергии. Печень способна расщеплять аминокислоты до углеродного скелета, которые затем могут быть включены в цикл лимонной кислоты. Это похоже на то, как глюкоза используется для производства АТФ. Если диета человека не содержит достаточного количества углеводов и жиров, его организм будет использовать больше аминокислот для производства энергии, что ставит под угрозу синтез новых белков и разрушает мышечные белки.Кроме того, если в рационе человека содержится больше белка, чем необходимо организму, лишние аминокислоты расщепляются и превращаются в жир.
Рисунок 6.14 Аминокислоты, используемые для производства энергии
Изображение Эллисон Калабрезе / CC BY 4.0
Электронное обучение — Белки
Брат Грегори говорит со своим классом,
На сегодняшний день предметом обсуждения является структура и функция белков .Вы должны следовать уроку, отвечать на вопросы, а затем, при необходимости, завершить исследовательское расследование.
«Начнем …….
«Белки — одна из четырех основных групп макромолекул, которые встречаются во всех живых организмах. Эти гигантские молекулы выполняют многие жизненно важные функции, необходимые клеткам.
«Белки участвуют в таких процессах, как переваривание пищи, структура клеток, катализ, движение, манипуляции с энергией и многое другое.Это сложные огромные ассоциации молекулярных субъединиц, которые невозможно понять. К счастью, все они построены по одному и тому же принципу.
«Как и все макромолекулы, белки представляют собой полимеры, состоящие из более мелких субъединиц — аминокислот, соединенных вместе в длинные цепи.
«В большинстве белков содержится около 20-22 общих аминокислот. Все эти небольшие молекулы, кроме одной, имеют одинаковую общую структуру, но различаются по природе одной химической группы — называемой« R-группа ».Это различная структура и свойства этих R-групп , которые делают аминокислоты отличными друг от друга.
«Аминокислоты объединены в длинные цепи, называемые« полипептиды », название происходит от типа связи, удерживающей цепи аминокислот вместе. Группа атомов, которые удерживают аминокислоты вместе, называется пептидной связью • Порядок или последовательность аминокислот в полипептидной цепи устанавливает первое критическое свойство белков, их первичную структуру.
Продолжая этот урок, проверьте себя, отвечая на некоторые из этих вопросов типа «правда / ложь».
Трехмерные формы
«Большинство свойств большинства белков, однако, основаны на уникальных и особых трехмерных формах, которые белки принимают, когда аминокислоты действуют и реагируют друг с другом и с окружающей водой.Вторичные, третичные и четвертичные структуры — это разные уровни более высокой трехмерной структуры.
«Форма очень важна. Аминокислота R-группы могут быть сгруппированы в четыре основные категории, и именно взаимодействие этих различных R-групп в значительной степени определяет форму и роль конечных белок.
«В большинстве белков имеется различное количество вторичной структуры, но два наиболее распространенных, узнаваемых повторяющихся паттерна структуры — это альфа-спираль и бета-складчатый лист.
«Хотя именно пептидная связь соединяет аминокислоты вместе в их полипептидных цепях, существуют другие, более слабые силы, удерживающие полипептид в его правильной трехмерной форме.
«Эти силы могут« сваривать »две части цепи вместе или просто обеспечивать локальную гидрофобную среду, из которой исключены молекулы воды.
«Обретя форму, белковые молекулы способны объединяться и образовывать сложные комплексы или структуры, которые состоят из более чем одной полипептидной субъединицы.Довольно часто эти более крупные структуры также содержат дополнительный небелковый материал, такой как углеводы, липиды или даже полинуклеотиды.
Roles Proteins Play — несколько примеров
«Белки участвуют почти во всех аспектах клеточной жизни и структуры. Они также играют важную роль в удерживании клеток вместе в многоклеточные структуры (как мы!) И обеспечивают протекание химических реакций с« жизненной скоростью ».
«Ниже приводится частичный список с несколькими примерами ролей, которые белки играют в жизни:
Научно-исследовательское исследование
проверьте свое расписание, чтобы узнать, требуется ли это
Проклятие Амона Исследование белка Исследование
Вопросы концепции
к уроку
проверьте свое расписание, чтобы узнать, требуется ли это
Protein Set Concept Вопросы и страница личных вопросов
Функция белка всегда зависит от встречи двух или более молекул: субстрат-фермент, лиганд-рецептор и т.д. теплового перемешивания. Учитывая величину и дальнодействующий характер электростатических взаимодействий, неудивительно, что они могут играть определяющую роль в этом процессе, управляя и / или ориентируя входящую молекулу [1].В случае ферментов дополнительным функциональным требованием является стабилизация промежуточного продукта реакции, без которого они не могут действовать как катализаторы. Также здесь большое значение имеют электростатические взаимодействия, при этом остатки активного центра стабилизируют промежуточное распределение заряда [2]. Гораздо более важным условием функционирования белка является наличие правильно сложенной структуры. Гидрофобный эффект, который обычно считается наиболее определяющим фактором в общем процессе сворачивания, по существу является следствием того факта, что полярные и особенно заряженные остатки электростатически более стабильны при взаимодействии с водой, чем с более неполярной внутренней частью белка.Кроме того, водородные связи, ответственные за вторичную структуру белка, в основном являются результатом электростатических взаимодействий. Таким образом, электростатические взаимодействия занимают особое место среди факторов, определяющих функцию и стабильность белков, и их правильное понимание и моделирование имеют большое значение. Источниками электростатических взаимодействий в белках являются заряженные титруемые остатки (Asp, Glu, His, Lys, Arg, C- и N-концы, Tyr и свободный Cys), структурные ионы (например, Mg 2 + в PGM), связанные ионы, противоионы раствора и др.В следующем разделе мы представляем некоторые модели, которые можно использовать для вычисления электростатических взаимодействий, возникающих из-за этих зарядов, и для связи их с функциональными характеристиками белков. Общие обзоры электростатики белков: Warshel & Russel (1984) [3], Matthew (1985) [4], Rogers (1986) [5], Harvey (1989) [6], Sharp & Honig (1990) [7], Дэвис и Маккаммон (1990) [1].
Аспект, который практически невозможно отделить от белковой электростатики, — это влияние pH из-за изменчивости заряда титруемых остатков.Иногда драматическая функциональная зависимость функции белка от pH является результатом протонирования и депротонирования этих остатков, что может привести к выраженным изменениям во взаимодействии электростатических взаимодействий. Поначалу теоретическая задача объяснения и предсказания таких электростатических изменений при pH может показаться простой — с учетом pK a титруемых остатков (доступных в любом справочнике по биохимии) было бы тривиальным вопросом сказать, какие остатки заряжены при данном pH.К сожалению, ситуация намного сложнее, потому что, как объясняется ниже, pK a может быть сдвинуто на несколько единиц pH от своего типичного значения. Фактически, даже обычная концепция pK a становится неуместной. Таким образом, чтобы решить, какой заряд должен быть отнесен к остатку при заданном pH, необходимо прибегнуть к довольно более сложным методам, как обсуждается ниже.
Протеом органелл — Атлас белков человека
Пространственное разделение биологических процессов — это фундаментальное явление для жизни, которое позволяет нескольким процессам происходить параллельно без нежелательного вмешательства.Органелла — это субъединица эукариотической клетки со специальной функцией. Название «органелла» происходит от аналогии между различными ролями органелл в клетках и различными ролями органов в организме человека в целом. Часто различают органеллы, связанные с мембраной, и органеллы, не связанные с мембраной. Связанные с мембраной органеллы, такие как ядро и аппарат Гольджи, имеют четко определенную физическую границу, которая разделяет внутри- и внеорганическое пространство. Напротив, органеллы, не связанные с мембраной, такие как цитоскелет и ядрышки, представляют собой пространственно различные сборки белков, а иногда и РНК внутри клетки.Мембранно или нет, это разделение создает особую среду в месте расположения органеллы, где концентрация различных молекул может быть адаптирована в соответствии с назначением органеллы, и предоставляет важную возможность для регуляции клеточных процессов. Поскольку точное определение органелл варьируется, более инклюзивные термины субклеточной структуры.
Основная функция белков — катализировать, проводить и контролировать клеточные процессы во времени и пространстве. Поскольку разные органеллы и субклеточные структуры предлагают различные среды, с различными физиологическими условиями и партнерами по взаимодействию, субклеточная локализация белка является важной частью функции белка.Следовательно, неправильная локализация белков часто была связана с клеточной дисфункцией и различными заболеваниями человека (Kau TR et al. (2004); Laurila K et al. (2009); Park S et al. (2011)). Знание пространственного распределения белка на субклеточном уровне важно для понимания функции белка, взаимодействий и клеточных механизмов, а изучение того, как клетки генерируют и поддерживают свою пространственную организацию, является центральным для понимания механизмов живых клеток.
В клеточном атласе субклеточная локализация 12813 белков была сопоставлена на одноклеточном уровне с 35 субклеточными структурами, что позволило определить 13 основных протеомов органелл.Анализ показывает, что примерно половина белков локализуется в нескольких компартментах, и идентифицирует многие белки с одноклеточными вариациями с точки зрения изобилия или пространственного распределения. Паттерн экспрессии и пространственное распределение белков человека во всех основных клеточных органеллах можно изучить в этих интерактивных разделах знаний, которые включают многочисленные каталоги белков со специфическими и схожими паттернами экспрессии, а также примеры изображений, иллюстрирующие различные паттерны внутриклеточного пространственного распределения.
Субклеточная локализация белков
Описано несколько подходов к систематическому анализу локализации белков. Количественные масс-спектрометрические данные позволяют идентифицировать белки со сходными профилями распределения по градиентам фракционирования (Park S et al. (2011); Christoforou A et al. (2016); Itzhak DN et al. (2016)) или ферментно-опосредованную близость. меченые белки в клетках (Itzhak DN et al. (2016); Roux KJ et al. (2012); Lee SY et al. (2016)). Напротив, подходы, основанные на визуализации, позволяют исследовать субклеточное распределение белков in situ в отдельных клетках и имеют преимущество в том, что они эффективно идентифицируют изменчивость отдельных клеток и локализацию в нескольких органеллах.Подходы на основе визуализации могут быть выполнены с использованием меченых белков (Huh WK et al. (2003); Simpson JC et al. (2000); Stadler C et al. (2013)) или аффинных реагентов.
В Cell Atlas мы используем подход, основанный на иммунофлуоресценции (IF), в сочетании с конфокальной микроскопией, чтобы обеспечить исследование пространственного распределения белков с высоким разрешением (Thul PJ et al. (2017); Stadler C et al. (2013); Барбе Л. и др. (2008); Штадлер С. и др. (2010); Фагерберг Л. и др. (2011)). С ограниченным дифракцией разрешением около 200 нм иммунофлуоресцентное изображение из Cell Atlas дает подробное представление о клеточной организации.Пространственное распределение белка исследуют с использованием непрямого IF в клеточной линии U-2 OS и до двух дополнительных клеточных линий, выбранных на основе экспрессии мРНК соответствующего гена, с использованием панели из 35 клеточных линий. Интересующий белок отображается зеленым цветом, а контрольные маркеры для микротрубочек (красный), эндоплазматического ретикулума (желтый) и ядра (синий) используются для обозначения клетки. От маленьких точек, таких как ядерные тельца, до более крупных структур, таких как нуклеоплазма, различные узоры на изображениях вместе с контрольными маркерами позволяют точно определить пространственное распределение белка в клетке.Локализация каждого белка назначается одной или нескольким из 35 субклеточных структур и субструктур, которые в настоящее время аннотированы в Cell Atlas, как показано на рисунке 1.
Нуклеоплазма
Ядерные крапинки
Ядерные тела
Ядрышки
Фибриллярный центр ядрышек
Ободок ядрышек
Митотическая хромосома
Кинетохора
Ядерная мембрана
Цитозоль
Цитоплазматические тела
Жезлы и Кольца
Агрессивный
Митохондрии
Центросома
Центриолярные спутники
Микротрубочки
Концы микротрубочек
Митотическое веретено
Цитокинетический мост
Midbody
Кольцо Midbody
Борозда спайности
Промежуточные волокна
Актиновые нити
Места очагового спайки
Эндоплазматическая сеть
аппарат Гольджи
Везикулы
Эндосомы
Лизосомы
Липидные капли
Пероксисомы
Плазматическая мембрана
Клеточные соединения
Рисунок 1.Пример конфокальных иммунофлуоресцентных изображений различных белков (зеленый цвет), локализованных в каждой из субклеточных органелл и субструктур, аннотированных в настоящее время в Атласе клеток в репрезентативном наборе клеточных линий. Микротрубочки помечаются антителом к тубулину (красный), а ядро окрашивается DAPI (синий). Сторона изображения составляет 64 мкм. Для получения дополнительных примеров изображений и деталей, описывающих все 35 шаблонов, аннотированных в Cell Atlas, см. Cell Dictionary.
Распределение белка в клетке человека
Рисунок 2 показывает распределение органелл всех аннотаций для белков 12813, локализованных по крайней мере в одной структуре или субструктуре.Участок отсортирован по мета-компартментам: цитоплазма, ядро и секреторный аппарат соответственно. Большинство белков находится в ядре, за ними следуют цитозоль и везикулы, которые состоят из транспортных пузырьков, а также небольших мембраносвязанных органелл, таких как эндосомы или пероксисомы. 55% (n = 7 · 106) белков были обнаружены более чем в одном месте (мультилокализирующие белки), а 25% (n = 3141) показали одноклеточную вариацию уровня экспрессии или пространственного распределения.
Рисунок 2.Гистограмма, показывающая распределение белков, обнаруженных в каждой органелле, структуре и субструктуре, аннотированных в Атласе клеток.
Проверка антител и данных о местоположении для Cell Atlas
Качество и использование антител в исследованиях часто обсуждаются (Baker M & period; (2015)). Поскольку связывание антитела с мишенью может привести к ложноположительным результатам, Cell Atlas пытается вручную подсчитать все результаты относительно надежности окрашивания. В Атласе клеток представлена оценка надежности для каждого аннотированного местоположения по четырехбалльной шкале: улучшенная, поддерживаемая, одобренная и неопределенная, как подробно описано в разделе «Анализ и аннотации».Улучшенные местоположения получены посредством валидации антител в соответствии с одним из «столпов» валидации, предложенных международной рабочей группой (Uhlen M et al. (2016): (i) генетические методы с использованием сайленсинга siRNA (Stadler C et al. (2012) ) или нокаут CRISPR / Cas9, (ii) экспрессия флуоресцентного белка, меченного белком, на эндогенных уровнях (Skogs M et al. (2017)) или (iii) независимые антитела, нацеленные на разные эпитопы (Stadler C et al. (2010) Поддерживаемое местоположение соответствует внешним экспериментальным данным (база данных UniProt), в то время как оценка утвержденного местоположения указывает на отсутствие внешней экспериментальной информации для подтверждения наблюдаемого местоположения.Неопределенное местоположение противоречит дополнительной информации, такой как данные литературы или транскриптомики, и показано, если нельзя исключить, что данные верны, и необходимы дальнейшие эксперименты для установления надежности окрашивания антител. Распределение оценок надежности для локализованных белков показано на рисунке 3. Приблизительно 43% (n = 5502) предоставленных локализаций белков улучшены или поддерживаются. В таблице 1 подробно описано распределение всех локализованных белков в органеллах и распределение оценок надежности на основе отдельных органелл.
Рис. 3. Круговая диаграмма, показывающая уровень надежности локализованных белков, где каждая часть представляет собой количество белков с одним типом оценки из четырех оценок надежности: Повышенная, Поддерживаемая, Одобренная и Неопределенная.
Таблица 1. Таблица, показывающая количество белков, локализованных в каждой органелле, структуре и субструктуре в Атласе клеток, а также распределение оценок надежности.
Соответствующие ссылки и публикации
Парих К. и др., Разнообразие эпителиальных клеток толстой кишки при здоровье и воспалительном заболевании кишечника и периоде; Природа и период; (2019) PubMed: 30814735 DOI: 10.1038 / s41586-019-0992-y
Menon M et al., Одноклеточный транскриптомный атлас сетчатки глаза человека определяет типы клеток, связанные с возрастной дегенерацией желтого пятна & period; Нац Коммуна & период; (2019) PubMed: 31653841 DOI: 10.1038 / s41467-019-12780-8
Wang L et al., Одноклеточная реконструкция сердца взрослого человека во время сердечной недостаточности и восстановления выявляет клеточный ландшафт, лежащий в основе сердечной функции и периода; Nat Cell Biol & period; (2020) PubMed: 313 DOI: 10.1038 / s41556-019-0446-7
Wang Y et al., Анализ одноклеточного транскриптома показывает различные функции всасывания питательных веществ в кишечнике человека & период; J Exp Med & period; (2020) PubMed: 31753849 DOI: 10.1084 / jem.201
Liao J et al., Секвенирование одноклеточной РНК почек человека & период; Научные данные и период; (2020) PubMed: 31896769 DOI: 10.1038 / s41597-019-0351-8
MacParland SA et al., Секвенирование одноклеточной РНК печени человека выявляет различные внутрипеченочные популяции макрофагов & период; Нац Коммуна & период; (2018) PubMed: 30348985 DOI: 10.1038 / s41467-018-06318-7
Vieira Braga FA et al., Перепись клеток легких человека выявляет новые состояния клеток при здоровье и астме & периоде; Nat Med & period; (2019) PubMed: 31209336 DOI: 10.1038 / s41591-019-0468-5
Vento-Tormo R et al., Одноклеточная реконструкция раннего взаимодействия матери и плода у людей и период; Природа и период; (2018) PubMed: 30429548 DOI: 10.1038 / s41586-018-0698-6
Qadir MMF et al., Анализ одноклеточного разрешения ниши клеток-предшественников протоков поджелудочной железы человека & период; Proc Natl Acad Sci U S A & period; (2020) PubMed: 32354994 DOI: 10.1073 / pnas.14117
Solé-Boldo L et al., Одноклеточные транскриптомы кожи человека показывают возрастную потерю прайминга фибробластов & период; Commun Biol & period; (2020) PubMed: 32327715 DOI: 10.1038 / s42003-020-0922-4
Henry GH et al., Клеточная анатомия простаты и уретры простаты взрослого человека & period; Cell Rep & period; (2018) PubMed: 30566875 DOI: 10.1016 / j.celrep.2018.11.086
Chen J et al., Фиксация и обработка PBMC для секвенирования одноклеточной РНК хрома & период; J Transl Med & period; (2018) PubMed: 30016977 DOI: 10.1186 / s12967-018-1578-4
Guo J et al., Атлас транскрипционных клеток семенников взрослого человека. Cell Res. (2018) PubMed: 30315278 DOI: 10.1038 / s41422-018-0099-2
Uhlen M. et al., Предложение по валидации антител. Нат. Методы. (2016) PubMed: 27595404 DOI: 10.1038 / nmeth.3995
Stadler C et al., Систематическая проверка связывания антител и субклеточной локализации белков с использованием миРНК и конфокальной микроскопии. Дж. Протеомика. (2012) PubMed: 22361696 DOI: 10.1016 / j.jprot.2012.01.030
Poser I et al., BAC TransgeneOmics & Colon; высокопроизводительный метод исследования функции белков у млекопитающих и периода; Nat Методы и период; (2008) PubMed: 183 DOI: 10.1038 / nmeth.1199
Skogs M et al., Валидация антител в приложениях биоимиджинга на основе эндогенной экспрессии меченых белков. J Proteome Res. (2017) PubMed: 27723985 DOI: 10.1021 / acs.jproteome.6b00821
Takahashi H et al., 5 ‘центрированное по концам профили экспрессии с использованием экспрессии генов cap-analysis и секвенирования следующего поколения & period; Nat Protoc & period; (2012) PubMed: 22362160 DOI: 10.1038 / nprot.2012.005
Lein ES et al., Полногеномный атлас экспрессии генов в мозге взрослой мыши & period; Природа и период; (2007) PubMed: 17151600 DOI: 10.1038 / nature05453
Kircher M et al., Двойное индексирование устраняет неточности в мультиплексном секвенировании на платформе Illumina и период; Nucleic Acids Res & period; (2012) PubMed: 22021376 DOI: 10.1093 / nar / gkr771
Pollard TD et al., Actin & comma; центральный игрок в форме клетки, движении и периоде; Наука и период; (2009) PubMed: 19965462 DOI: 10.1126 / science.1175862
Mitchison TJ et al., Подвижность клеток на основе актина и перемещение клеток и период; Ячейка и период; (1996) PubMed: 8608590
Pollard TD et al., Молекулярный механизм цитокинеза & период; Annu Rev Biochem & period; (2019) PubMed: 30649923 DOI: 10.1146 / annurev-biochem-062917-012530
dos Remedios CG et al., Актин-связывающие белки и толстая кишка; регуляция цитоскелетных микрофиламентов и периода; Physiol Rev & period; (2003) PubMed: 12663865 DOI: 10.1152 / Physrev.00026.2002
Campellone KG et al., Гонка вооружений нуклеаторов и двоеточие; клеточный контроль сборки актина и периода; Nat Rev Mol Cell Biol & period; (2010) PubMed: 20237478 DOI: 10.1038 / nrm2867
Rottner K et al., Краткий обзор механизмов сборки актина и период; J Cell Sci & period; (2017) PubMed: 2
57 DOI: 10.1242 / jcs.206433
Bird RP & period ;, Наблюдение и количественная оценка аберрантных крипт в толстой кишке мыши, обработанной канцерогеном толстой кишки & col; предварительные выводы и период; Рак Lett & period; (1987) PubMed: 3677050 DOI: 10.1016 / 0304-3835 (87) -1
HUXLEY AF et al., Структурные изменения в мышцах во время сокращения & semi; интерференционная микроскопия живых мышечных волокон и периода; Природа и период; (1954) PubMed: 13165697
HUXLEY H et al., Изменения поперечных полос мышц во время сокращения и растяжения и их структурная интерпретация и период; Природа и период; (1954) PubMed: 13165698
Svitkina T & period ;, Актиновый цитоскелет и подвижность на основе актина & период; Cold Spring Harb Perspect Biol & period; (2018) PubMed: 29295889 DOI: 10.1101 / cshperspect.a018267
Kelpsch DJ et al., Nuclear Actin & Colon; От открытия до функции и периода; Анат Рек & lpar; Хобокен & rpar; & period; (2018) PubMed: 30312531 DOI: 10.1002 / ar.23959
Malumbres M et al., Клеточный цикл и запятая; CDK и рак и толстая кишка; меняющаяся парадигма и период; Nat Rev Рак и период; (2009) PubMed: 1
48 DOI: 10.1038 / nrc2602
Massagué J & period ;, G1 контроль клеточного цикла и рак & период; Природа и период; (2004) PubMed: 15549091 DOI: 10.1038 / nature03094
Hartwell LH et al., Контроль клеточного цикла и рак & период; Наука и период; (1994) PubMed: 7997877 DOI: 10.1126 / science.7997877
Barnum KJ et al., Регулирование клеточного цикла с помощью контрольных точек и периода; Методы Mol Biol & period; (2014) PubMed: 24
7 DOI: 10.1007 / 978-1-4939-0888-2_2
Weinberg RA & period ;, Белок ретинобластомы и контроль клеточного цикла & период; Ячейка и период; (1995) PubMed: 7736585 DOI: 10.1016 / 0092-8674 (95) -2
Morgan DO & period ;, Принципы регулирования CDK и период; Природа и период; (1995) PubMed: 7877684 DOI: 10.1038 / 374131a0
Teixeira LK et al., Убиквитин-лигазы и контроль клеточного цикла & период; Annu Rev Biochem & period; (2013) PubMed: 23495935 DOI: 10.1146 / annurev-biochem-060410-105307
King RW et al., Как протеолиз управляет клеточным циклом и периодом; Наука и период; (1996) PubMed: 8939846 DOI: 10.1126 / science.274.5293.1652
Cho RJ et al., Регуляция транскрипции и функция во время клеточного цикла человека & период; Nat Genet & period; (2001) PubMed: 11137997 DOI: 10.1038 / 83751
Whitfield ML et al., Идентификация генов, периодически экспрессируемых в клеточном цикле человека, и их экспрессия в опухолях & период; Mol Biol Cell & period; (2002) PubMed: 12058064 DOI: 10.1091 / mbc.02-02-0030.
Boström J et al., Сравнительная транскриптомика клеточного цикла показывает синхронизацию сетей онтогенетических факторов транскрипции в раковых клетках. PLoS One. (2017) PubMed: 202 DOI: 10.1371 / journal.pone.0188772
Lane KR et al., Изменения содержания белка, регулируемого клеточным циклом, в синхронно пролиферирующих клетках HeLa включают регуляцию белков сплайсинга пре-мРНК & период; PLoS One & period; (2013) PubMed: 23520512 DOI: 10.1371 / journal.pone.0058456
Ohta S et al., Белковый состав митотических хромосом, определенный с использованием мультиклассификатора комбинаторной протеомики и периода; Ячейка и период; (2010) PubMed: 20813266 DOI: 10.1016 / j.cell.2010.07.047
Ly T et al., Протеомная хронология экспрессии генов через клеточный цикл в клетках миелоидного лейкоза человека & период; Элиф и период; (2014) PubMed: 24596151 DOI: 10.7554 / eLife.01630
Pagliuca FW et al., Количественная протеомика раскрывает основу биохимической специфичности механизма и периода клеточного цикла; Mol Cell & period; (2011) PubMed: 21816347 DOI: 10.1016 / j.molcel.2011.05.031
Ly T et al., Протеомный анализ ответа на остановку клеточного цикла в клетках миелоидного лейкоза человека & период; Элиф и период; (2015) PubMed: 25555159 DOI: 10.7554 / eLife.04534
Dueck H et al., Вариация — функция & двоеточие; Функционально ли важны различия в отдельных клетках & quest; & col; Проверка гипотезы о том, что для агрегированной функции & period; Биологические исследования и период; (2016) PubMed: 26625861 DOI: 10.1002 / bies.201500124
Thul PJ et al., Субклеточная карта протеома человека. Наука. (2017) PubMed: 28495876 DOI: 10.1126 / science.aal3321
Cooper S et al., Анализ содержания циклинов A & comma посредством мембранного элюирования; B1 & запятая; и E во время невозмущенного клеточного цикла & period; Ячейка Div & период; (2007) PubMed: 17892542 DOI: 10.1186 / 1747-1028-2-28
Davis PK et al., Биологические методы синхронизации клеточного цикла клеток млекопитающих & период; Биотехнологии и период; (2001) PubMed: 11414226 DOI: 10.2144 / 01306rv01
Domenighetti G et al., Влияние информационной кампании в СМИ на частоту и период гистерэктомии; Ланцет и период; (1988) PubMed: 2
1 DOI: 10.1016 / s0140-6736 (88) -9
Scialdone A et al., Вычислительное отнесение стадии клеточного цикла к данным транскриптома одиночной клетки & период; Методы и период; (2015) PubMed: 26142758 DOI: 10.1016 / j.ymeth.2015.06.021
Sakaue-Sawano A et al., Визуализация пространственно-временной динамики развития многоклеточного клеточного цикла и периода; Ячейка и период; (2008) PubMed: 18267078 DOI: 10.1016 / j.cell.2007.12.033
Grant GD et al., Идентификация генов, регулируемых клеточным циклом, периодически экспрессируемых в клетках U2OS, и их регуляция факторами транскрипции FOXM1 и E2F & period; Mol Biol Cell & period; (2013) PubMed: 24109597 DOI: 10.1091 / mbc.E13-05-0264
Semple JW et al., Существенная роль Orc6 в репликации ДНК посредством поддержания пререпликативных комплексов & период; EMBO J & период; (2006) PubMed: 17053779 DOI: 10.1038 / sj.emboj.7601391
Kilfoil ML et al., Стохастическая вариация и двоеточие; от одиночных клеток до суперорганизмов & период; HFSP J & период; (2009) PubMed: 20514130 DOI: 10.2976 / 1.3223356
Ansel J et al., Стохастическая изменчивость экспрессии генов от клетки к клетке является сложным генетическим признаком & период; PLoS Genet & period; (2008) PubMed: 18404214 DOI: 10.1371 / journal.pgen.1000049
Colman-Lerner A et al., Регулируемая межклеточная изменчивость в системе решения клеточной судьбы & период; Природа и период; (2005) PubMed: 16170311 DOI: 10.1038 / nature03998
Liberali P et al., Одноклеточные и многомерные подходы к скринингу генетических нарушений и период; Nat Rev Genet & period; (2015) PubMed: 25446316 DOI: 10.1038 / nrg3768
Elowitz MB et al., Стохастическая экспрессия гена в одной клетке & период; Наука и период; (2002) PubMed: 12183631 DOI: 10.1126 / science.1070919
Kaern M et al., Стохастичность экспрессии генов и толстой кишки; от теорий к фенотипам и периоду; Nat Rev Genet & period; (2005) PubMed: 15883588 DOI: 10.1038 / nrg1615
Bianconi E et al., Оценка количества клеток в организме человека & период; Ann Hum Biol & period; (2013) PubMed: 23829164 DOI: 10.3109 / 03014460.2013.807878
Collins K et al., Клеточный цикл и рак & период; Proc Natl Acad Sci U S A & period; (1997) PubMed: 91
Животовский Б. и др., Клеточный цикл и гибель клеток при заболевании и толстой кишке; прошедшее & запятая; настоящее и будущее и период; J Intern Med & period; (2010) PubMed: 20964732 DOI: 10.1111 / j.1365-2796.2010.02282.x
Cho RJ et al., Полногеномный транскрипционный анализ митотического клеточного цикла и периода; Mol Cell & period; (1998) PubMed: 9702192
Spellman PT et al., Всесторонняя идентификация регулируемых клеточным циклом генов дрожжей Saccharomyces cerevisiae с помощью гибридизации на микрочипах и период; Mol Biol Cell & period; (1998) PubMed: 9843569
Orlando DA et al., Глобальный контроль транскрипции клеточного цикла с помощью связанных генераторов CDK и сети & period; Природа и период; (2008) PubMed: 18463633 DOI: 10.1038 / nature06955
Rustici G et al., Программа периодической экспрессии генов клеточного цикла делящихся дрожжей & период; Nat Genet & period; (2004) PubMed: 15195092 DOI: 10.1038 / ng1377
Uhlén M. et al., Тканевая карта протеома человека. Наука (2015) PubMed: 25613900 DOI: 10.1126 / science.1260419
Nigg EA et al., Цикл центросомы и толстая кишка; Биогенез центриолей & запятая; дублирование и врожденная асимметрия и период; Nat Cell Biol & period; (2011) PubMed: 21968988 DOI: 10.1038 / ncb2345
Doxsey S & period ;, Повторная оценка функции и периода центросомы; Nat Rev Mol Cell Biol & period; (2001) PubMed: 11533726 DOI: 10.1038 / 35089575
Bornens M & period ;, Состав центросом и механизмы закрепления микротрубочек и период; Curr Opin Cell Biol & period; (2002) PubMed: 11792541
Conduit PT et al., Функция и сборка центросом в клетках животных & период; Nat Rev Mol Cell Biol & period; (2015) PubMed: 26373263 DOI: 10.1038 / nrm4062
Tollenaere MA et al., Центриолярные спутники и толстая кишка; ключевые медиаторы функций и периода центросом; Cell Mol Life Sci & period; (2015) PubMed: 25173771 DOI: 10.1007 / s00018-014-1711-3
Prosser SL et al., Центриолярный сателлитный биогенез и функция в клетках позвоночных и период; J Cell Sci & period; (2020) PubMed: 31896603 DOI: 10.1242 / jcs.239566
Rieder CL et al., Центросома позвоночных и толстой кишки; больше, чем центр организации микротрубочек и период; Trends Cell Biol & period; (2001) PubMed: 11567874
Badano JL et al., Центросома в генетических заболеваниях человека & период; Nat Rev Genet & period; (2005) PubMed: 15738963 DOI: 10.1038 / nrg1557
Clegg JS & period ;, Свойства и метаболизм водной цитоплазмы и ее границы & период; Am J Physiol & period; (1984) PubMed: 6364846
Luby-Phelps K & period ;, Физическая химия цитоплазмы и ее влияние на функцию клеток & толстой кишки; обновление & период; Mol Biol Cell & period; (2013) PubMed: 23989722 DOI: 10.1091 / mbc.E12-08-0617
Luby-Phelps K & period ;, Цитоархитектура и физические свойства цитоплазмы и толстой кишки; объем и запятая; вязкость и запятая; диффузия и запятая; площадь и период внутриклеточной поверхности; Int Rev Cytol & period; (2000) PubMed: 10553280
Ellis RJ & period ;, Макромолекулярное скопление и толстая кишка; очевидный, но недооцененный & период; Trends Biochem Sci & period; (2001) PubMed: 115
Bright GR et al., Визуализирующая микроскопия с соотношением флуоресценции и толстой кишки; временные и пространственные измерения цитоплазматического pH и периода; J Cell Biol & period; (1987) PubMed: 3558476
Kopito RR & period ;, Aggresomes & comma; тельца включения и агрегация белков и период; Trends Cell Biol & period; (2000) PubMed: 11121744
Aizer A et al., Внутриклеточное перемещение и динамика P-телец и период; Прион и период; (2008) PubMed: 193
Carcamo WC et al., Молекулярная клеточная биология и иммунобиология стержневых и зольчатых структур и периода у млекопитающих; Int Rev Cell Mol Biol & period; (2014) PubMed: 24411169 DOI: 10.1016 / B978-0-12-800097-7.00002-6
Lang F & period ;, Механизмы и значение регулирования объема клеток & период; J Am Coll Nutr & period; (2007) PubMed: 174
Schwarz DS et al., Эндоплазматический ретикулум и толстая кишка; структура и запятая; функция и ответ на сотовую сигнализацию & период; Cell Mol Life Sci & period; (2016) PubMed: 26433683 DOI: 10.1007 / s00018-015-2052-6
Friedman JR et al., ER в 3D и двоеточие; многофункциональная динамическая мембранная сеть & период; Trends Cell Biol & period; (2011) PubMed: 21
9 DOI: 10.1016 / j.tcb.2011.07.004
Travers KJ et al., Функциональный и геномный анализ выявляет существенную координацию между развернутым белковым ответом и ER-ассоциированной деградацией & период; Ячейка и период; (2000) PubMed: 10847680
Roussel BD et al., Дисфункция эндоплазматического ретикулума при неврологических заболеваниях и периоде; Lancet Neurol & period; (2013) PubMed: 23237905 DOI: 10.1016 / S1474-4422 (12) 70238-7
Neve EP et al., Белки цитохрома P450 и толстая кишка; удержание и распространение из эндоплазматической сети и периода; Curr Opin Drug Discov Devel & period; (2010) PubMed: 20047148
Kulkarni-Gosavi P et al., Форма и функция аппарата Гольджи и толстой кишки; строительные леса и запятая; цитоскелет и передача сигналов и период; FEBS Lett & period; (2019) PubMed: 31378930 DOI: 10.1002 / 1873-3468.13567
Short B et al., Аппарат Гольджи и период; Curr Biol & period; (2000) PubMed: 10985372 DOI: 10.1016 / s0960-9822 (00) 00644-8
Wei JH et al., Распутывая ленту Гольджи и период; Трафик и период; (2010) PubMed: 21040294 DOI: 10.1111 / j.1600-0854.2010.01114.x
Wilson C et al., Аппарат Гольджи и толстая кишка; органелла с множеством сложных функций & период; Biochem J & period; (2011) PubMed: 21158737 DOI: 10.1042 / BJ20101058
Farquhar MG et al., Аппарат Гольджи и толстая кишка; 100 лет прогресса, противоречий и периода; Trends Cell Biol & period; (1998) PubMed: 9695800
Brandizzi F et al., Организация интерфейса ER-Golgi для мембранного управления трафиком & период; Nat Rev Mol Cell Biol & period; (2013) PubMed: 23698585 DOI: 10.1038 / nrm3588
Potelle S et al., Посттрансляционные модификации Гольджи и связанные с ними заболевания и период; J Наследовать Metab Dis & period; (2015) PubMed: 25967285 DOI: 10.1007 / s10545-015-9851-7
Leduc C et al., Промежуточные филаменты в миграции и инвазии клеток & толстой кишки; необычные подозреваемые и период; Curr Opin Cell Biol & period; (2015) PubMed: 25660489 DOI: 10.1016 / j.ceb.2015.01.005
Лоури Дж. И др., Промежуточные волокна играют ключевую роль в регулировании архитектуры и функции клеток & период; J Biol Chem & period; (2015) PubMed: 25957409 DOI: 10.1074 / jbc.R115.640359
Роберт А. и др., Динамика промежуточных волокон и толстой кишки; Что мы видим сейчас и почему это важно & period; Биологические исследования и период; (2016) PubMed: 26763143 DOI: 10.1002 / bies.201500142
Fuchs E et al., Промежуточные волокна и толстая кишка; структура и запятая; динамика и запятая; функция & запятая; и болезнь и период; Annu Rev Biochem & period; (1994) PubMed: 7979242 DOI: 10.1146 / annurev.bi.63.070194.002021
Janmey PA et al., Вязкоупругие свойства виментина по сравнению с другими нитевидными биополимерными сетками & период; J Cell Biol & period; (1991) PubMed: 2007620
Köster S et al., Механика промежуточных волокон in vitro и в клетке и толстой кишке; от спиральных катушек до нитей и запятой; волокна и сети и период; Curr Opin Cell Biol & period; (2015) PubMed: 25621895 DOI: 10.1016 / j.ceb.2015.01.001
Herrmann H et al., Промежуточные волокна и толстая кишка; от клеточной архитектуры до наномеханики и периода; Nat Rev Mol Cell Biol & period; (2007) PubMed: 17551517 DOI: 10.1038 / nrm2197
Gauster M et al., Кератины в трофобласте человека и период; Histol Histopathol & period; (2013) PubMed: 23450430 DOI: 10.14670 / HH-28.817
Janke C & period ;, Код тубулина и двоеточие; молекулярные компоненты и запятая; механизмы считывания и запятая; и функции & период; J Cell Biol & period; (2014) PubMed: 25135932 DOI: 10.1083 / jcb.201406055
Goodson HV et al., Микротрубочки и ассоциированные с микротрубочками белки & period; Cold Spring Harb Perspect Biol & period; (2018) PubMed: 29858272 DOI: 10.1101 / cshperspect.a022608
Wade RH & period ;, На и вокруг микротрубочек и толстой кишки; обзор и период; Mol Biotechnol & period; (2009) PubMed: 19565362 DOI: 10.1007 / s12033-009-9193-5
Desai A et al., Динамика и период полимеризации микротрубочек; Annu Rev Cell Dev Biol & period; (1997) PubMed: 9442869 DOI: 10.1146 / annurev.cellbio.13.1.83
Conde C et al., Сборка микротрубочек и запятая; организация и динамика в аксонах и дендритах и периоде; Nat Rev Neurosci & period; (2009) PubMed: 19377501 DOI: 10.1038 / nrn2631
Schmoranzer J et al., Роль микротрубочек в слиянии пост-везикул Гольджи с плазматической мембраной & period; Mol Biol Cell & period; (2003) PubMed: 12686609 DOI: 10.1091 / mbc.E02-08-0500
Skop AR et al., Диссекция протеома среднего тела млекопитающего выявляет механизмы и период консервативного цитокинеза; Наука и период; (2004) PubMed: 15166316 DOI: 10.1126 / science.1097931
Waters AM et al., Цилиопатии и толстая кишка; расширяющийся спектр болезней и период; Педиатр Нефрол и период; (2011) PubMed: 21210154 DOI: 10.1007 / s00467-010-1731-7
Matamoros AJ et al., Микротрубочки в здоровье и дегенеративных заболеваниях нервной системы & период; Brain Res Bull & период; (2016) PubMed: 27365230 DOI: 10.1016 / j.brainresbull.2016.06.016
Jordan MA et al., Микротрубочки как мишень для противоопухолевых препаратов и период; Nat Rev Рак и период; (2004) PubMed: 15057285 DOI: 10.1038 / nrc1317
Nunnari J et al., Митохондрии и толстая кишка; в болезни и в здоровье и периоде; Ячейка и период; (2012) PubMed: 22424226 DOI: 10.1016 / j.cell.2012.02.035
Friedman JR et al., Форма и функция митохондрий и период; Природа и период; (2014) PubMed: 24429632 DOI: 10.1038 / nature12985
Calvo SE et al., Митохондриальный протеом и болезнь человека и период; Annu Rev Genomics Hum Genet & period; (2010) PubMed: 206
DOI: 10.1146 / annurev-genom-082509-141720
McBride HM et al., Митохондрии и толстая кишка; больше, чем просто электростанция и период; Curr Biol & period; (2006) PubMed: 16860735 DOI: 10.1016 / j.cub.2006.06.054
Schaefer AM et al., Эпидемиология митохондриальных нарушений — прошлое & запятая; настоящее и будущее и период; Biochim Biophys Acta & period; (2004) PubMed: 15576042 DOI: 10.1016 / j.bbabio.2004.09.005
Lange A et al., Классические сигналы ядерной локализации & двоеточие; определение & запятая; функция & запятая; и взаимодействие с importin alpha & period; J Biol Chem & period; (2007) PubMed: 17170104 DOI: 10.1074 / jbc.R600026200
Ашмарина Л.И. и др., 3-гидрокси-3-метилглутарил-кофермент А лиаза & толстая кишка; нацеливание и обработка в пероксисомах и митохондриях & период; J Lipid Res & period; (1999) PubMed: 9869651
Wang SC et al., Ядерная транслокация рецепторов тирозинкиназы мембранных рецепторов эпидермального фактора роста & период; Clin Cancer Res & period; (2009) PubMed: 19861462 DOI: 10.1158 / 1078-0432.CCR-08-2813
Jeffery CJ & period ;, Moonlighting protein & period; Trends Biochem Sci & period; (1999) PubMed: 10087914
Jeffery CJ & period ;, Зачем изучать подрабатывающие белки и поиски; Спереди Genet & period; (2015) PubMed: 26150826 DOI: 10.3389 / fgene.2015.00211
Pancholi V & period ;, Многофункциональная альфа-енолаза & двоеточие; его роль в болезнях и периоде; Cell Mol Life Sci & period; (2001) PubMed: 11497239 DOI: 10.1007 / pl00000910
Chapple CE et al., Экстремальные многофункциональные белки, идентифицированные из сети взаимодействия белков человека & период; Нац Коммуна & период; (2015) PubMed: 26054620 DOI: 10.1038 / ncomms8412
Dechat T. et al., Ядерные ламины и толстая кишка; основные факторы структурной организации и функции ядра и хроматина & период; Genes Dev & period; (2008) PubMed: 18381888 DOI: 10.1101 / gad.1652708
Gruenbaum Y et al., Ядерная пластинка достигает возраста и периода; Nat Rev Mol Cell Biol & period; (2005) PubMed: 15688064 DOI: 10.1038 / nrm1550
Stuurman N. et al., Ядерные ламины и толстая кишка; их структура и запятая; сборка и запятая; и взаимодействия & период; J Struct Biol & period; (1998) PubMed: 9724605 DOI: 10.1006 / jsbi.1998.3987
Paine PL et al., Проницаемость ядерной оболочки и период; Природа и период; (1975) PubMed: 1117994
Reichelt R et al., Корреляция между структурой и массовым распределением ядерного порового комплекса и отдельных компонентов порового комплекса & период; J Cell Biol & period; (1990) PubMed: 2324201
CALLAN HG et al., Экспериментальные исследования ядер и периодов ооцитов амфибий; Я & период; Исследование структуры ядерной мембраны с помощью электронного микроскопа & period; Proc R Soc Lond B Biol Sci & period; (1950) PubMed: 14786306
WATSON ML & period ;, Ядерная оболочка & semi; его структура и отношение к цитоплазматическим мембранам & период; J Biophys Biochem Cytol & period; (1955) PubMed: 13242591
BAHR GF et al., Тонкая структура ядерной мембраны личиночной слюнной железы и средней кишки Chironomus & period; Exp Cell Res & period; (1954) PubMed: 13173504
Terasaki M et al., Новая модель разрушения ядерной оболочки и периода; Mol Biol Cell & period; (2001) PubMed: 11179431
Dultz E et al., Систематический кинетический анализ митотической разборки и повторной сборки ядерной поры в живых клетках & период; J Cell Biol & period; (2008) PubMed: 18316408 DOI: 10.1083 / jcb.200707026
Salina D et al., Цитоплазматический динеин как средство разрушения ядерной оболочки и период; Ячейка и период; (2002) PubMed: 117
Beaudouin J et al., Разрушение ядерной оболочки происходит за счет вызванного микротрубочками разрыва пластинки & период; Ячейка и период; (2002) PubMed: 117
Gerace L et al., Пластинка ядерной оболочки обратимо деполимеризуется во время митоза & период; Ячейка и период; (1980) PubMed: 7357605
Ellenberg J et al., Динамика ядерной мембраны и повторная сборка в живых клетках & толстой кишке; нацеливание на белок внутренней ядерной мембраны в интерфазе и митозе & периоде; J Cell Biol & period; (1997) PubMed: 9298976
Yang L et al., Интегральные мембранные белки ядерной оболочки рассредоточены по эндоплазматическому ретикулуму во время митоза & период; J Cell Biol & period; (1997) PubMed: 56
Bione S et al., Идентификация нового Х-сцепленного гена, ответственного за мышечную дистрофию Эмери-Дрейфуса и период; Nat Genet & period; (1994) PubMed: 7894480 DOI: 10.1038 / ng1294-323
Boisvert FM et al., Многофункциональное ядрышко и период; Nat Rev Mol Cell Biol & period; (2007) PubMed: 17519961 DOI: 10.1038 / nrm2184
Scheer U et al., Структура и функция ядрышка и период; Curr Opin Cell Biol & period; (1999) PubMed: 10395554 DOI: 10.1016 / S0955-0674 (99) 80054-4
Németh A et al., Организация генома в ядрышке и вокруг него & период; Тенденции Genet & period; (2011) PubMed: 21295884 DOI: 10.1016 / j.tig.2011.01.002
Cuylen S et al., Ki-67 действует как биологическое поверхностно-активное вещество для диспергирования митотических хромосом и периода; Природа и период; (2016) PubMed: 27362226 DOI: 10.1038 / nature18610
Stenström L et al., Картирование протеома ядрышка выявляет пространственно-временную организацию, связанную с внутренним нарушением белков. Mol Syst Biol. (2020) PubMed: 32744794 DOI: 10.15252 / msb.20209469
Derenzini M et al., Размер ядра указывает на скорость пролиферации клеток в раковых тканях & период; J Pathol & period; (2000) PubMed: 10861579 DOI: 10.1002 / (SICI) 1096-9896 (200006) 191: 2 <181 :: AID-PATH607> 3.0.CO; 2-V
Visintin R et al., Ядрышко и толстая кишка ; шляпа фокусника для фокусов клеточного цикла и периода; Curr Opin Cell Biol & period; (2000) PubMed: 10801456
Marciniak RA et al., Ядрышковая локализация белка синдрома Вернера в клетках человека & period; Proc Natl Acad Sci U S A & period; (1998) PubMed: 9618508
Tamanini F et al., Хрупкие X-родственные белки FXR1P и FXR2P содержат функциональный сигнал нацеливания на ядрышки, эквивалентный регуляторным белкам ВИЧ-1 & period; Hum Mol Genet & period; (2000) PubMed: 10888599
Willemsen R et al., Ассоциация FMRP с частицами-предшественниками рибосом в ядрышке & период; Biochem Biophys Res Commun & period; (1996) PubMed: 8769090 DOI: 10.1006 / bbrc.1996.1126
Isaac C et al., Характеристика продукта ядрышкового гена & запятая; патока и запятая; в синдроме Тричера Коллинза и периоде; Mol Biol Cell & period; (2000) PubMed: 10982400
Drygin D et al., Механизм транскрипции РНК-полимеразы I & coli; новая цель для лечения рака и периода; Annu Rev Pharmacol Toxicol & period; (2010) PubMed: 20055700 DOI: 10.1146 / annurev.pharmtox.010909.105844
Spector DL & period;, Макромолекулярные домены в ядре клетки & period; Annu Rev Cell Biol & period; (1993) PubMed: 8280462 DOI: 10.1146 / annurev.cb.09.110193.001405
Ламонд А.И. и др., Структура и функции ядра и периода; Наука и период; (1998) PubMed: 9554838
SWIFT H & period ;, Исследования ядерной тонкой структуры и периода; Brookhaven Symp Biol & period; (1959) PubMed: 13836127
Ламонд А.И. и др., Ядерные точки & толстая кишка; модель ядерных органелл и период; Nat Rev Mol Cell Biol & period; (2003) PubMed: 12
2 DOI: 10.1038 / nrm1172
Thiry M & period ;, Гранулы межхроматина & период; Histol Histopathol & period; (1995) PubMed: 8573995
Sleeman JE et al., Вновь собранные snRNP ассоциируются со свернутыми телами перед пятнами и запятой; предполагая ядерный путь созревания snRNP и период; Curr Biol & period; (1999) PubMed: 10531003
Darzacq X et al., малые ядерные РНК, специфичные для тельца Кахаля, & ободочная кишка; новый класс направляющих РНК 2′-O-метилирования и псевдоуридилирования & период; EMBO J & период; (2002) PubMed: 12032087 DOI: 10.1093 / emboj / 21.11.2746
Jády BE et al., Модификация малых ядерных РНК Sm происходит в нуклеоплазматическом теле Кахаля после импорта из цитоплазмы & period; EMBO J & период; (2003) PubMed: 12682020 DOI: 10.1093 / emboj / cdg187
Liu Q et al., Новая ядерная структура, содержащая белок & период выживания моторных нейронов; EMBO J & период; (1996) PubMed: 8670859
Lefebvre S et al., Идентификация и характеристика гена, определяющего спинальную мышечную атрофию & period; Ячейка и период; (1995) PubMed: 7813012
Fischer U et al., Комплекс SMN-SIP1 играет важную роль в биогенезе сплайсосомных snRNP & period; Ячейка и период; (1997) PubMed: 9323130
Lallemand-Breitenbach V et al., PML ядерные тела и период; Cold Spring Harb Perspect Biol & period; (2010) PubMed: 20452955 DOI: 10.1101 / cshperspect.a000661
Booth DG et al., Ki-67 и компартмент периферии хромосомы при митозе и периоде; Trends Cell Biol & period; (2017) PubMed: 28838621 DOI: 10.1016 / j.tcb.2017.08.001
Ljungberg O et al., Составная фолликулярно-парафолликулярная клеточная карцинома щитовидной железы и толстой кишки; новая опухоль и квест; Рак & период; (1983) PubMed: 6136320 DOI: 10.1002 / 1097-0142 (19830915) 52: 6 <1053 :: aid-cncr2820520621> 3.0.co; 2-q
Melcák I et al., Ядерная компартментализация пре-мРНК и толстая кишка; трафик выпущенных транскриптов в резервуары и периоды факторов сплайсинга; Mol Biol Cell & period; (2000) PubMed: 10679009
Spector DL et al., Связи между отдельными компонентами сплайсинга пре-мРНК и ядром клетки & period; EMBO J & период; (1991) PubMed: 1833187
Misteli T et al., Фосфорилирование белка и ядерная организация сплайсинга пре-мРНК & period; Trends Cell Biol & period; (1997) PubMed: 17708924 DOI: 10.1016 / S0962-8924 (96) 20043-1
Cmarko D et al., Ультраструктурный анализ транскрипции и сплайсинга в ядре клетки после микроинъекции бром-UTP & period; Mol Biol Cell & period; (1999) PubMed: 9880337
Van Hooser AA et al., Перихромосомный слой и период; Хромосома и период; (2005) PubMed: 16136320 DOI: 10.1007 / s00412-005-0021-9
Booth DG et al., Ki-67 представляет собой белок, взаимодействующий с PP1, который организует периферию митотической хромосомы & period; Элиф и период; (2014) PubMed: 24867636 DOI: 10.7554 / eLife.01641
Kau TR et al., Ядерный транспорт и рак и толстая кишка; от механизма к вмешательству и периоду; Nat Rev Рак и период; (2004) PubMed: 14732865 DOI: 10.1038 / nrc1274
Laurila K et al., Прогнозирование связанных с заболеванием мутаций, влияющих на локализацию и период белка; BMC Genomics & period; (2009) PubMed: 19309509 DOI: 10.1186 / 1471-2164-10-122
Park S. et al., Локализация белка как основной признак этиологии и коморбидности генетических заболеваний & период; Mol Syst Biol & period; (2011) PubMed: 21613983 DOI: 10.1038 / msb.2011.29
Christoforou A et al., Черновик карты пространственного протеома и периода плюрипотентных стволовых клеток мыши; Нац Коммуна & период; (2016) PubMed: 26754106 DOI: 10.1038 / ncomms9992
Itzhak DN et al., Global & comma; количественное и динамическое картирование субклеточной локализации и периода белка; Элиф и период; (2016) PubMed: 27278775 DOI: 10.7554 / eLife.16950
Roux KJ et al., Беспорядочный гибридный белок с биотин-лигазой идентифицирует проксимальные и взаимодействующие белки в клетках млекопитающих & period; J Cell Biol & period; (2012) PubMed: 22412018 DOI: 10.1083 / jcb.201112098
Lee SY et al., APEX Fingerprinting выявляет субклеточную локализацию интересующих белков & period; Cell Rep & period; (2016) PubMed: 27184847 DOI: 10.1016 / j.celrep.2016.04.064
Huh WK et al., Глобальный анализ локализации белка у почкующихся дрожжей и период; Природа и период; (2003) PubMed: 14562095 DOI: 10.1038 / nature02026
Simpson JC et al., Систематическая субклеточная локализация новых белков, идентифицированных с помощью крупномасштабного секвенирования кДНК & период; EMBO Rep & period; (2000) PubMed: 11256614 DOI: 10.1093 / embo-reports / kvd058
Stadler C et al., Иммунофлуоресценция и мечение флуоресцентным белком показывают высокую корреляцию для локализации белка в клетках млекопитающих. Нат. Методы. 2013 Apr; 10 (4): 315-23 (2013) PubMed: 23435261 DOI: 10.1038 / nmeth.2377
Barbe L. et al., К конфокальному субклеточному атласу протеома человека. Mol Cell Proteomics. (2008) PubMed: 18029348 DOI: 10.1074 / mcp.M700325-MCP200
Stadler C et al., Единый протокол фиксации для исследований локализации иммунофлуоресценции в масштабе протеома. Дж. Протеомика. (2010) PubMed: 19896565 DOI: 10.1016 / j.jprot.2009.10.012
Fagerberg L et al., Картирование субклеточного распределения белка в трех линиях клеток человека. J Proteome Res. (2011) PubMed: 21675716 DOI: 10.1021 / pr200379a
Baker M & period ;, Кризис воспроизводимости и двоеточие; Во всем виноваты антитела и период; Природа и период; (2015) PubMed: 25993940 DOI: 10.


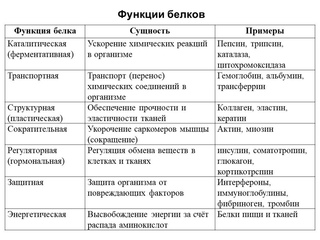
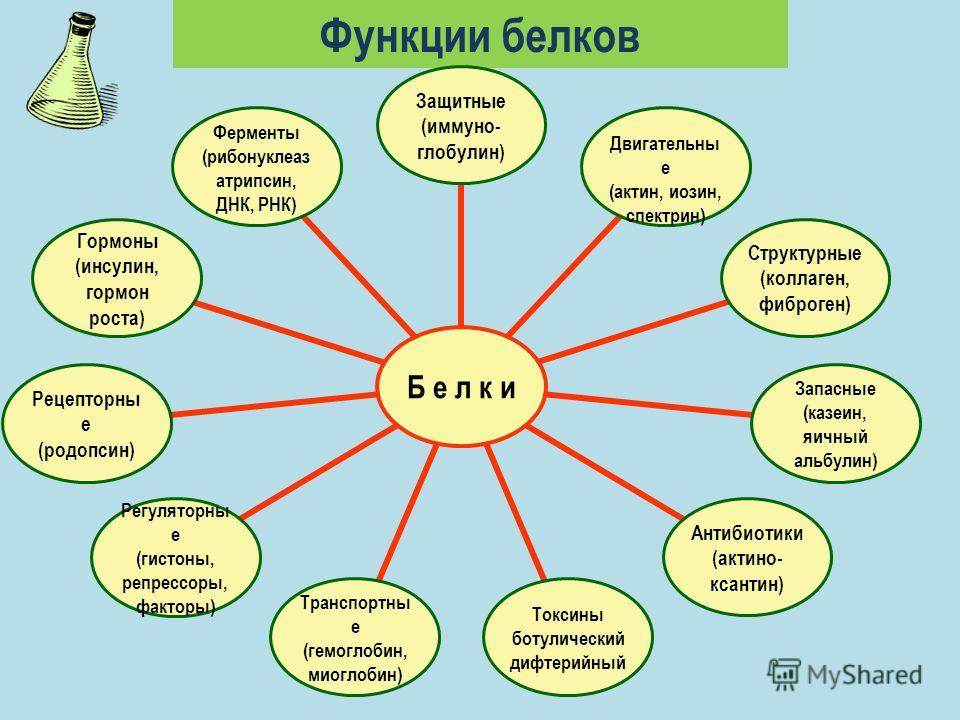 Цитохромы осуществляют перенос электронов по электронтранспортным цепям митохондрий и хлоропластов.
Цитохромы осуществляют перенос электронов по электронтранспортным цепям митохондрий и хлоропластов.  Белки входят в состав всех живых организмов. Мышцы, кости, покровные ткани, внутренние органы, хрящи, шерсть, кровь — все это белковые вещества.
Белки входят в состав всех живых организмов. Мышцы, кости, покровные ткани, внутренние органы, хрящи, шерсть, кровь — все это белковые вещества.
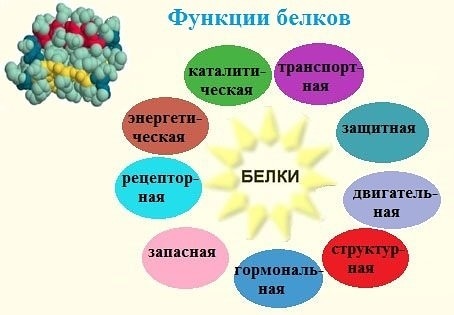






 Даже временное отсутствие одной незаменимой аминокислоты может отрицательно сказаться на синтезе белка. При уменьшении количества любой незаменимой аминокислоты или ее отсутствии пропорционально уменьшается эффективность всех остальных аминокислот.
Даже временное отсутствие одной незаменимой аминокислоты может отрицательно сказаться на синтезе белка. При уменьшении количества любой незаменимой аминокислоты или ее отсутствии пропорционально уменьшается эффективность всех остальных аминокислот.
 Ряд других белков несомненно используется в организме в качестве источника аминокислот, которые в свою очередь являются предшественниками биологически активных веществ, регулирующих процессы обмена веществ.
Ряд других белков несомненно используется в организме в качестве источника аминокислот, которые в свою очередь являются предшественниками биологически активных веществ, регулирующих процессы обмена веществ. В качестве другого примера защитной роли можно привести способность ряда белков крови к свертыванию. Свертывание белка плазмы крови фибриногена приводит к образованию сгустка крови, что предохраняет от потери крови при ранениях.
В качестве другого примера защитной роли можно привести способность ряда белков крови к свертыванию. Свертывание белка плазмы крови фибриногена приводит к образованию сгустка крови, что предохраняет от потери крови при ранениях. д. Наконец, в комплексе с липидами (в частности, фосфолипидами) белки участвуют в образовании биомембран клеток.
д. Наконец, в комплексе с липидами (в частности, фосфолипидами) белки участвуют в образовании биомембран клеток.
 д., а соединение, состоящее из 20 и более аминокислотных остатков, – полипептидом.
д., а соединение, состоящее из 20 и более аминокислотных остатков, – полипептидом.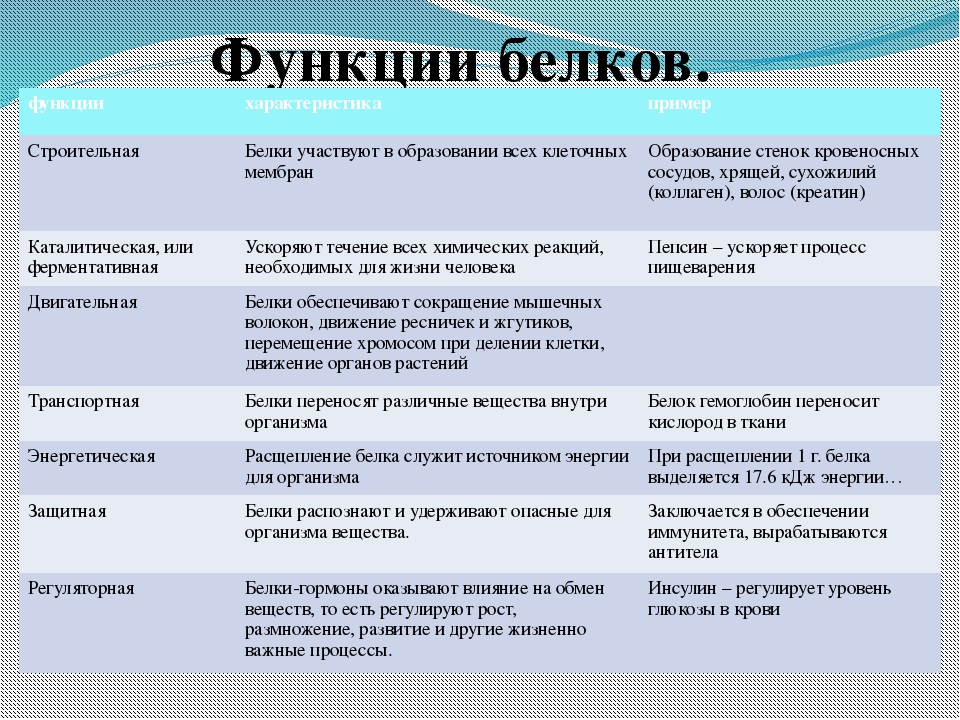 Атомы серы двух аминокислот, находящихся на некотором расстоянии друг от друга в полипептидной цепи, соединяются, образуя так называемые дисульфидные, или S–S, связи. Благодаря этим взаимодействиям, а также другим, менее сильным связям, белковая спираль сворачивается и приобретает форму шарика, или глобулы. Способ укладки полипептидных спиралей в глобуле называют третичной структурой белка. Многие белки, обладающие третичной структурой, могут выполнять свою биологическую роль в клетке. Однако для осуществления некоторых функций организма требуется участие белков с еще более высоким уровнем организации.
Атомы серы двух аминокислот, находящихся на некотором расстоянии друг от друга в полипептидной цепи, соединяются, образуя так называемые дисульфидные, или S–S, связи. Благодаря этим взаимодействиям, а также другим, менее сильным связям, белковая спираль сворачивается и приобретает форму шарика, или глобулы. Способ укладки полипептидных спиралей в глобуле называют третичной структурой белка. Многие белки, обладающие третичной структурой, могут выполнять свою биологическую роль в клетке. Однако для осуществления некоторых функций организма требуется участие белков с еще более высоким уровнем организации. В состав четвертичной структуры некоторых белков включаются помимо белковых субъединиц и разнообразные небелковые компоненты. Тот же гемоглобин содержит сложное гетероциклическое соединение, в состав которого входит железо.
В состав четвертичной структуры некоторых белков включаются помимо белковых субъединиц и разнообразные небелковые компоненты. Тот же гемоглобин содержит сложное гетероциклическое соединение, в состав которого входит железо.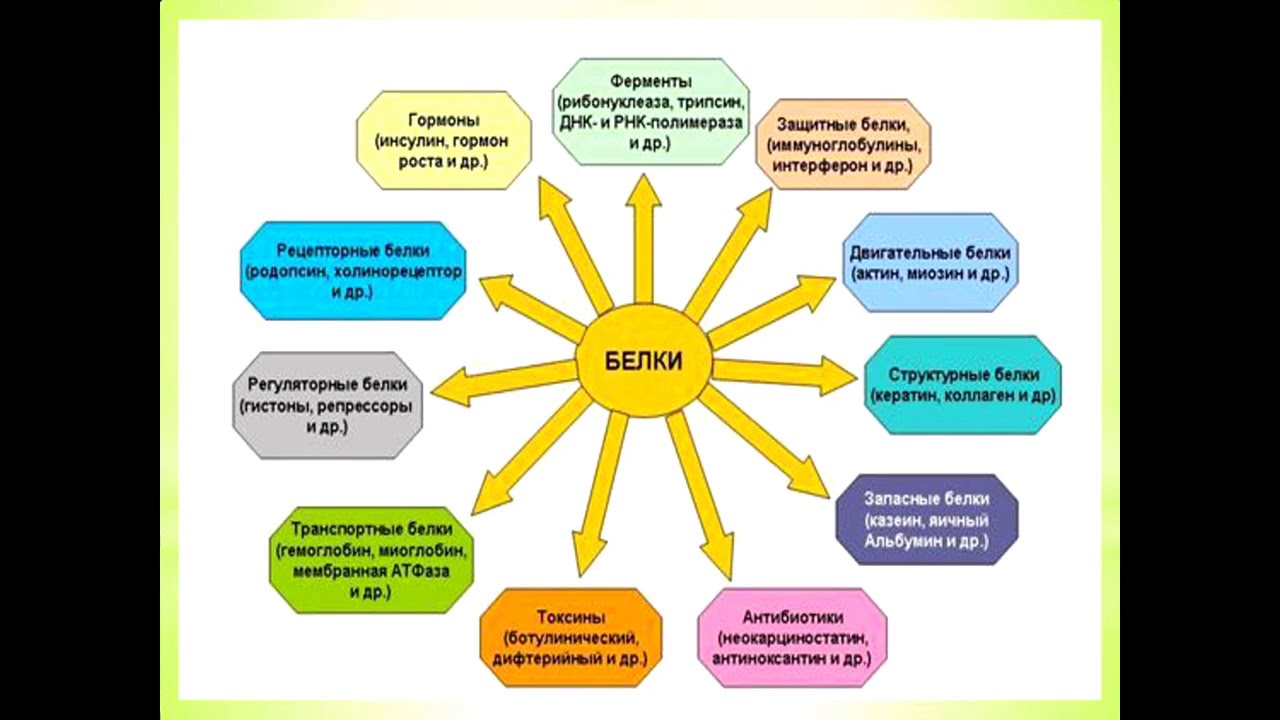
 Такой процесс носит название ренатурации.
Такой процесс носит название ренатурации.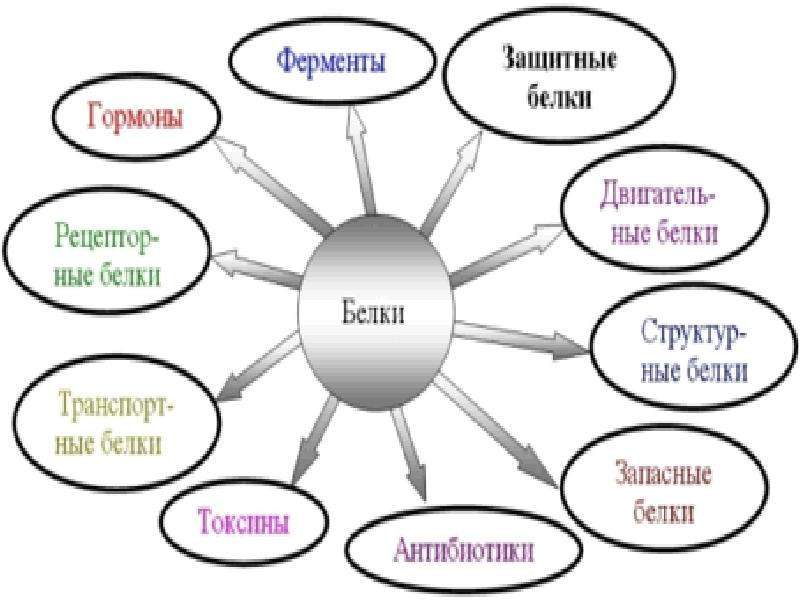 Так, в ответ на проникновение чужеродных объектов (антигенов) определенные лейкоциты вырабатывают специфические белки — иммуноглобулины (антитела), участвующие в иммунном ответе организма. Белок плазмы крови фибриноген, участвуя в свертывании крови и тем самым уменьшая кровопотери.
Так, в ответ на проникновение чужеродных объектов (антигенов) определенные лейкоциты вырабатывают специфические белки — иммуноглобулины (антитела), участвующие в иммунном ответе организма. Белок плазмы крови фибриноген, участвуя в свертывании крови и тем самым уменьшая кровопотери. В состав связок, стенок артерий и лёгких входит структурный белок эластин.
В состав связок, стенок артерий и лёгких входит структурный белок эластин.
Благодаря такому анализу после детального биохимического исследования мы некоторое время назад нашли новый взаимодействующий с каспазой-2 регуляторный фактор», – рассказал о работе руководитель научной группы и ведущий автор статьи Борис Животовский, доктор биологических наук и руководитель лаборатории исследования механизмов апоптоза факультета фундаментальной медицины МГУ имени М.В. Ломоносова.
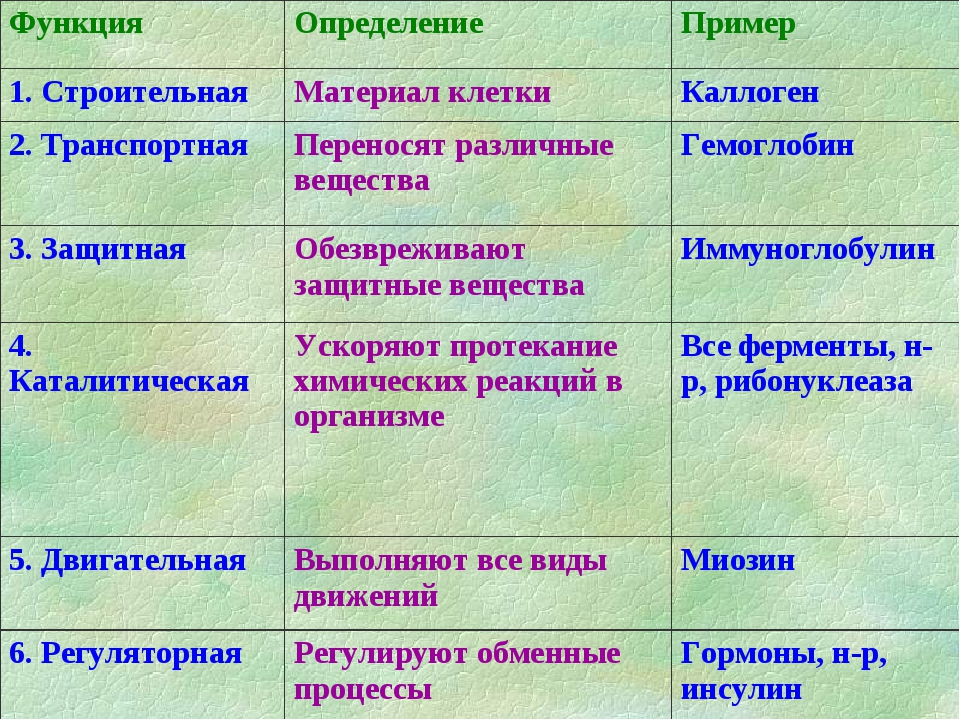 Кроме того, FAN контролирует миграцию клеток и производство интерлейкина-6 – молекулы, которая регулирует воспалительные процессы. Другая важная функция FAN – руководить образованием везикул, маленьких внутриклеточных пузырьков, состоящих из того же белково-липидного двухслойного «материала», что и клеточная мембрана. В таких пузырьках различные молекулы могут перемещаться по клетке, «перевариваться» с помощью специальных белков или выбрасываться наружу.
Кроме того, FAN контролирует миграцию клеток и производство интерлейкина-6 – молекулы, которая регулирует воспалительные процессы. Другая важная функция FAN – руководить образованием везикул, маленьких внутриклеточных пузырьков, состоящих из того же белково-липидного двухслойного «материала», что и клеточная мембрана. В таких пузырьках различные молекулы могут перемещаться по клетке, «перевариваться» с помощью специальных белков или выбрасываться наружу.– Последнее принципиально важно для понимания процесса метастазирования опухоли».
 Как мы помним, открытое в ходе этой же работы взаимодействие каспазы-2 с FAN влияет и на формирование везикул. Подавив действие каспазы-2, ученые убедились, что без этого белка внутриклеточные пузырьки сильно увеличивались в размере. Такая особенность везикул уже была ранее известна как проявление синдрома Чедиака-Хигаши, при котором у человека возникает светобоязнь, нервный тик и нарушения работы иммунной системы. Поэтому только дальнейшие исследования покажут, можно ли бороться с метастазами через подавление связывания каспазы-2 с FAN без вреда для других функций обоих белков.
Как мы помним, открытое в ходе этой же работы взаимодействие каспазы-2 с FAN влияет и на формирование везикул. Подавив действие каспазы-2, ученые убедились, что без этого белка внутриклеточные пузырьки сильно увеличивались в размере. Такая особенность везикул уже была ранее известна как проявление синдрома Чедиака-Хигаши, при котором у человека возникает светобоязнь, нервный тик и нарушения работы иммунной системы. Поэтому только дальнейшие исследования покажут, можно ли бороться с метастазами через подавление связывания каспазы-2 с FAN без вреда для других функций обоих белков. На первом
этапе в ходе беседы и рассказа учитель объясняет
функции белков, демонстрируя таблицы по общей
биологии. Объясняется значение каталазы в
растительной и животной клетке, ее функции –
расщепление опасно для клетки Н2О2 до
кислорода и воды.
На первом
этапе в ходе беседы и рассказа учитель объясняет
функции белков, демонстрируя таблицы по общей
биологии. Объясняется значение каталазы в
растительной и животной клетке, ее функции –
расщепление опасно для клетки Н2О2 до
кислорода и воды.

 Есть белки необычайно
активные (ферменты), есть химически неактивные,
устойчивые к воздействию различных условий
среды и крайне неустойчивые.
Есть белки необычайно
активные (ферменты), есть химически неактивные,
устойчивые к воздействию различных условий
среды и крайне неустойчивые.
 К запасным белкам относятся белки яйца,
белки мозга, белки сои.
К запасным белкам относятся белки яйца,
белки мозга, белки сои.
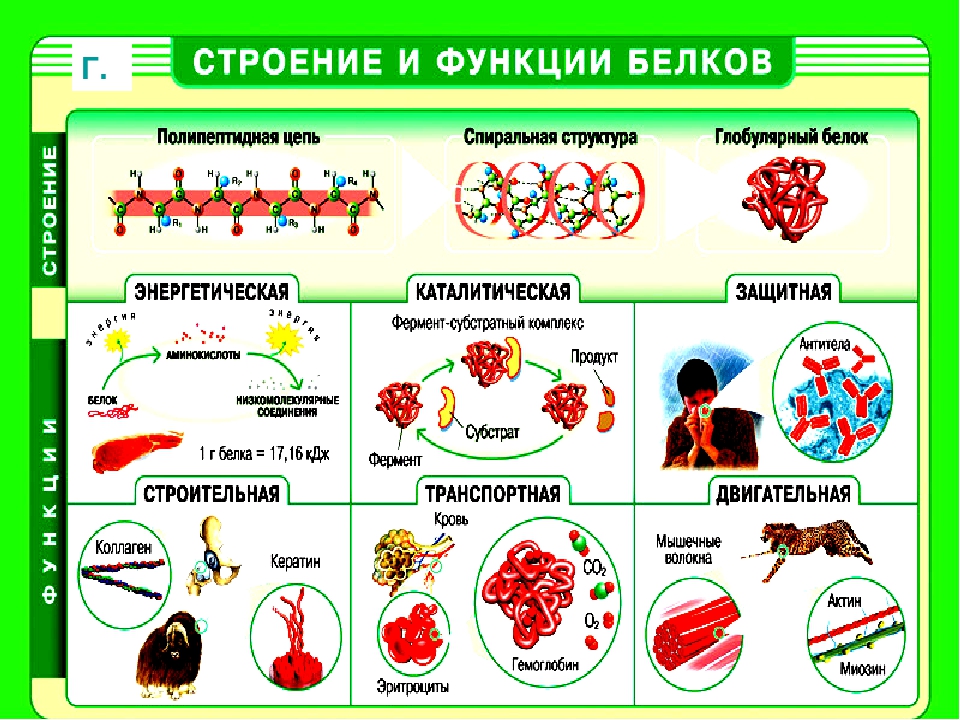 Объясните наблюдаемые вами явления
при действии Н2О2 на живые и мертвые
клетки (вареный картофель).
Объясните наблюдаемые вами явления
при действии Н2О2 на живые и мертвые
клетки (вареный картофель).
 профиль. Это может быть белок с измененной функцией или белок, который не функционирует — или даже сворачивается вообще »(Lesk, 2001).Например, гемоглобины взрослого человека и плода различаются заменой His на Ser (143β). В первую очередь, в результате этой мутации фетальный гемоглобин демонстрирует более низкое сродство, чем взрослый гомолог, к регуляторному лиганду дифосфоглицерату. Это способствует передаче кислорода плоду через плаценту (Lesk, 2001).
профиль. Это может быть белок с измененной функцией или белок, который не функционирует — или даже сворачивается вообще »(Lesk, 2001).Например, гемоглобины взрослого человека и плода различаются заменой His на Ser (143β). В первую очередь, в результате этой мутации фетальный гемоглобин демонстрирует более низкое сродство, чем взрослый гомолог, к регуляторному лиганду дифосфоглицерату. Это способствует передаче кислорода плоду через плаценту (Lesk, 2001).  2002; Ohno, 1970).Следующие примеры могут подтвердить это предположение. GroEL — это белок теплового шока 60 кДа, широко распространенный в бактериях. Интересно, что GroEL в Chlamydiae стал дублированным в происхождении линии Chlamydiae , представляющей три различных молекулярных шаперона, а именно исходный белок GroEL1 и его паралогичные белки GroEL2 и GroEL3 (Karunakaran et al. 2003; McNally and Fares, 2007 ). Виды копий белка GroEL функционально разошлись после событий дупликации генов.Функциональная дивергенция произошла в важных функциональных областях этих белков. Несмотря на то, что три белка GroEL Chlamidiae демонстрируют значительную консервацию аминокислотной последовательности в важных областях, участвующих в связывании полипептидов, по сравнению с GroEL из Escherichia coli , значительные различия были обнаружены в областях связывания GroEL и в областях, участвующих в связывании АТФ и гидролиз. Большинство аминокислотных замен, которые повлияли на взаимодействие с белками-партнерами и были ответственны за функциональную дивергенцию между паралогами GroEL, были исправлены адаптивной эволюцией после событий дупликации гена groEL (McNally and Fares, 2007).
2002; Ohno, 1970).Следующие примеры могут подтвердить это предположение. GroEL — это белок теплового шока 60 кДа, широко распространенный в бактериях. Интересно, что GroEL в Chlamydiae стал дублированным в происхождении линии Chlamydiae , представляющей три различных молекулярных шаперона, а именно исходный белок GroEL1 и его паралогичные белки GroEL2 и GroEL3 (Karunakaran et al. 2003; McNally and Fares, 2007 ). Виды копий белка GroEL функционально разошлись после событий дупликации генов.Функциональная дивергенция произошла в важных функциональных областях этих белков. Несмотря на то, что три белка GroEL Chlamidiae демонстрируют значительную консервацию аминокислотной последовательности в важных областях, участвующих в связывании полипептидов, по сравнению с GroEL из Escherichia coli , значительные различия были обнаружены в областях связывания GroEL и в областях, участвующих в связывании АТФ и гидролиз. Большинство аминокислотных замен, которые повлияли на взаимодействие с белками-партнерами и были ответственны за функциональную дивергенцию между паралогами GroEL, были исправлены адаптивной эволюцией после событий дупликации гена groEL (McNally and Fares, 2007).
 После дупликации молекулы STIM-1 и STIM-2, по-видимому, подверглись очищающей селекции. Кроме того, анализ последовательности связывающего домена Ca 2+ EF-hand и домена SAM вместе с функциональными исследованиями выявил критические области / остатки, которые, вероятно, лежат в основе функциональных изменений, и подтвердил доказательства гипотезы о том, что STIM-1 и STIM-2 могут развили отличные функциональные свойства после дупликации гена (Cai, 2007).
После дупликации молекулы STIM-1 и STIM-2, по-видимому, подверглись очищающей селекции. Кроме того, анализ последовательности связывающего домена Ca 2+ EF-hand и домена SAM вместе с функциональными исследованиями выявил критические области / остатки, которые, вероятно, лежат в основе функциональных изменений, и подтвердил доказательства гипотезы о том, что STIM-1 и STIM-2 могут развили отличные функциональные свойства после дупликации гена (Cai, 2007).  2006)].
2006)].  Эти типы событий склейки, вероятно, не будут представлены в коллекциях EST (Graveley, 2001). Некоторые гены производят несколько сплайсированных мРНК, в то время как оказывается, что некоторые другие гены фактически кодируют транскрипты, которые альтернативно сплайсируются для получения десятков тысяч различных мРНК (Schmucker et al.2000).
Эти типы событий склейки, вероятно, не будут представлены в коллекциях EST (Graveley, 2001). Некоторые гены производят несколько сплайсированных мРНК, в то время как оказывается, что некоторые другие гены фактически кодируют транскрипты, которые альтернативно сплайсируются для получения десятков тысяч различных мРНК (Schmucker et al.2000).  1995; Sugita et al. 1999). Кроме того, разнообразие белков нейроксинов может определять нейронные связи (Missler et al. 1998). Было показано, что взаимодействия β-нейрексинов, присутствующих в пресинаптических клетках, с нейролигинами на поверхности постсинаптических клеток достаточно для запуска образования синапсов (Scheiffele et al. 2000). Важно отметить, что это взаимодействие происходит только в том случае, если β-neurexin кодируется мРНК, лишенной альтернативного экзона 20.Белки, синтезированные из транскриптов, содержащих экзон 20, не взаимодействуют с нейролигинами (Ichtchenko et al. 1995). Таким образом, альтернативный сплайсинг транскриптов нейрексина может играть прямую роль в контроле образования и поддержания синапсов.
1995; Sugita et al. 1999). Кроме того, разнообразие белков нейроксинов может определять нейронные связи (Missler et al. 1998). Было показано, что взаимодействия β-нейрексинов, присутствующих в пресинаптических клетках, с нейролигинами на поверхности постсинаптических клеток достаточно для запуска образования синапсов (Scheiffele et al. 2000). Важно отметить, что это взаимодействие происходит только в том случае, если β-neurexin кодируется мРНК, лишенной альтернативного экзона 20.Белки, синтезированные из транскриптов, содержащих экзон 20, не взаимодействуют с нейролигинами (Ichtchenko et al. 1995). Таким образом, альтернативный сплайсинг транскриптов нейрексина может играть прямую роль в контроле образования и поддержания синапсов.  Как мы уже подчеркивали, на уровне генома максимальный объем генетической информации должен быть закодирован в минимальном пространстве ДНК. На уровне транскриптома скрытое разнообразие будущих белков, кодируемых в ДНК и мРНК, выявляется перетасовкой различных фрагментов первичного транскрипта во время альтернативного процесса сплайсинга. Использование альтернативного промотора, в свою очередь, раскрывает разнообразие генетической информации, содержащейся в мРНК и предназначенной для трансляции. На уровне протеома максимум структурных и функциональных потенциалов, содержащихся в генетической информации, встроенной в мРНК, может быть вызван трансляционными механизмами.Последние данные, полученные путем секвенирования генома человека, показали, что геном человека состоит из примерно 35 000 генов, которые могут экспрессировать примерно 100 000 белков. Такой выброс белка возможен только потому, что существуют многочисленные механизмы, с помощью которых функционально разнообразные полипептиды продуцируются из одного гена (Touriol et al.
Как мы уже подчеркивали, на уровне генома максимальный объем генетической информации должен быть закодирован в минимальном пространстве ДНК. На уровне транскриптома скрытое разнообразие будущих белков, кодируемых в ДНК и мРНК, выявляется перетасовкой различных фрагментов первичного транскрипта во время альтернативного процесса сплайсинга. Использование альтернативного промотора, в свою очередь, раскрывает разнообразие генетической информации, содержащейся в мРНК и предназначенной для трансляции. На уровне протеома максимум структурных и функциональных потенциалов, содержащихся в генетической информации, встроенной в мРНК, может быть вызван трансляционными механизмами.Последние данные, полученные путем секвенирования генома человека, показали, что геном человека состоит из примерно 35 000 генов, которые могут экспрессировать примерно 100 000 белков. Такой выброс белка возможен только потому, что существуют многочисленные механизмы, с помощью которых функционально разнообразные полипептиды продуцируются из одного гена (Touriol et al. 2003).
2003).  Канонические механизмы сканирования () управляют инициацией большинства мРНК, но также описаны три неклассических механизма инициации, зависящих от кэп: протекающее сканирование, рибосомное шунтирование и инициация терминации (Lopez-Lastra et al.2005).
Канонические механизмы сканирования () управляют инициацией большинства мРНК, но также описаны три неклассических механизма инициации, зависящих от кэп: протекающее сканирование, рибосомное шунтирование и инициация терминации (Lopez-Lastra et al.2005).  , 2003).
, 2003). 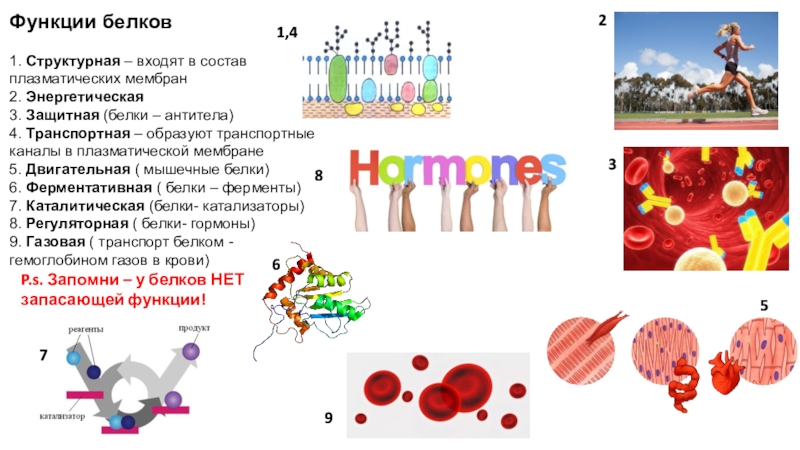 2006; Gandre и др., 2003 г .; Пакхэм и др., 1997 г.). Cai et al. (Cai et al. 2006) показали, что 85.7% событий альтернативной трансляции генерируют биологическое разнообразие, приписываемое различным субклеточным локализациям и содержанию различных доменов в альтернативных изоформах.Несколько примеров, представленных ниже, могут проиллюстрировать, как механизм сканирования с утечкой используется клеткой для увеличения разнообразия изоформ белка, продуцируемых одним геном.
2006; Gandre и др., 2003 г .; Пакхэм и др., 1997 г.). Cai et al. (Cai et al. 2006) показали, что 85.7% событий альтернативной трансляции генерируют биологическое разнообразие, приписываемое различным субклеточным локализациям и содержанию различных доменов в альтернативных изоформах.Несколько примеров, представленных ниже, могут проиллюстрировать, как механизм сканирования с утечкой используется клеткой для увеличения разнообразия изоформ белка, продуцируемых одним геном.  Инициация трансляции со второго стартового кодона в кадре дает изоформу AtLIG1, нацеленную только на ядро. Длина 5′-UTR и, что более важно, нуклеотидный контекст вокруг альтернативных стартовых кодонов в транскриптах AtLIG1 влияют на инициацию трансляции для обеспечения сбалансированного синтеза ядерных и митохондриальных изоформ AtLIG1 через контекстно-зависимый механизм сканирования дырявых рибосом (Sunderland et al. .2006 г.). Вместо того, чтобы иметь отдельные гены для кодирования различных изоформ белков ДНК-лигазы, специфичных для компартментов, эукариотические виды, от дрожжей до человека, по-видимому, используют эволюционно законсервированный механизм, который основан на выборе стартовых кодонов инициации трансляции в рамке считывания внутри рамки считывания транскрипта мРНК лигазы. для регулирования синтеза соответствующей изоформы ДНК-лигазы, предназначенной для ядра или митохондрий. Поскольку контроль субклеточной локализации белков и их взаимодействия со специфическими белками-партнерами in vivo имеют решающее значение для физиологии клетки, важность механизмов, контролирующих производство различных форм белка и их внутриклеточную топологию, трудно переоценить (Sunderland et al.
Инициация трансляции со второго стартового кодона в кадре дает изоформу AtLIG1, нацеленную только на ядро. Длина 5′-UTR и, что более важно, нуклеотидный контекст вокруг альтернативных стартовых кодонов в транскриптах AtLIG1 влияют на инициацию трансляции для обеспечения сбалансированного синтеза ядерных и митохондриальных изоформ AtLIG1 через контекстно-зависимый механизм сканирования дырявых рибосом (Sunderland et al. .2006 г.). Вместо того, чтобы иметь отдельные гены для кодирования различных изоформ белков ДНК-лигазы, специфичных для компартментов, эукариотические виды, от дрожжей до человека, по-видимому, используют эволюционно законсервированный механизм, который основан на выборе стартовых кодонов инициации трансляции в рамке считывания внутри рамки считывания транскрипта мРНК лигазы. для регулирования синтеза соответствующей изоформы ДНК-лигазы, предназначенной для ядра или митохондрий. Поскольку контроль субклеточной локализации белков и их взаимодействия со специфическими белками-партнерами in vivo имеют решающее значение для физиологии клетки, важность механизмов, контролирующих производство различных форм белка и их внутриклеточную топологию, трудно переоценить (Sunderland et al. 2006 г.).
2006 г.). 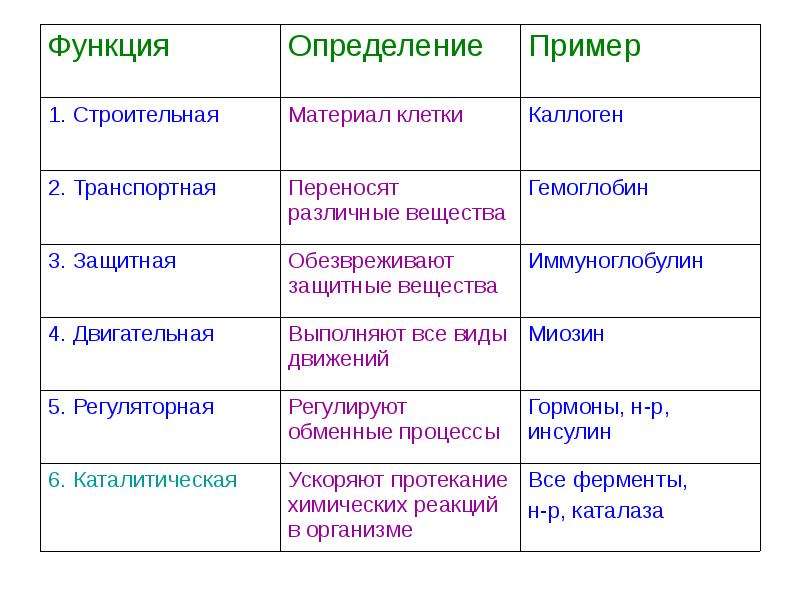 2006).
2006).  2006; Sass et al. 2001) демонстрируют силу механизмов просачивающегося сканирования в генерации множества функционально различных изоформ белков из одного транскрипта мРНК.
2006; Sass et al. 2001) демонстрируют силу механизмов просачивающегося сканирования в генерации множества функционально различных изоформ белков из одного транскрипта мРНК.