2. Формы поведения
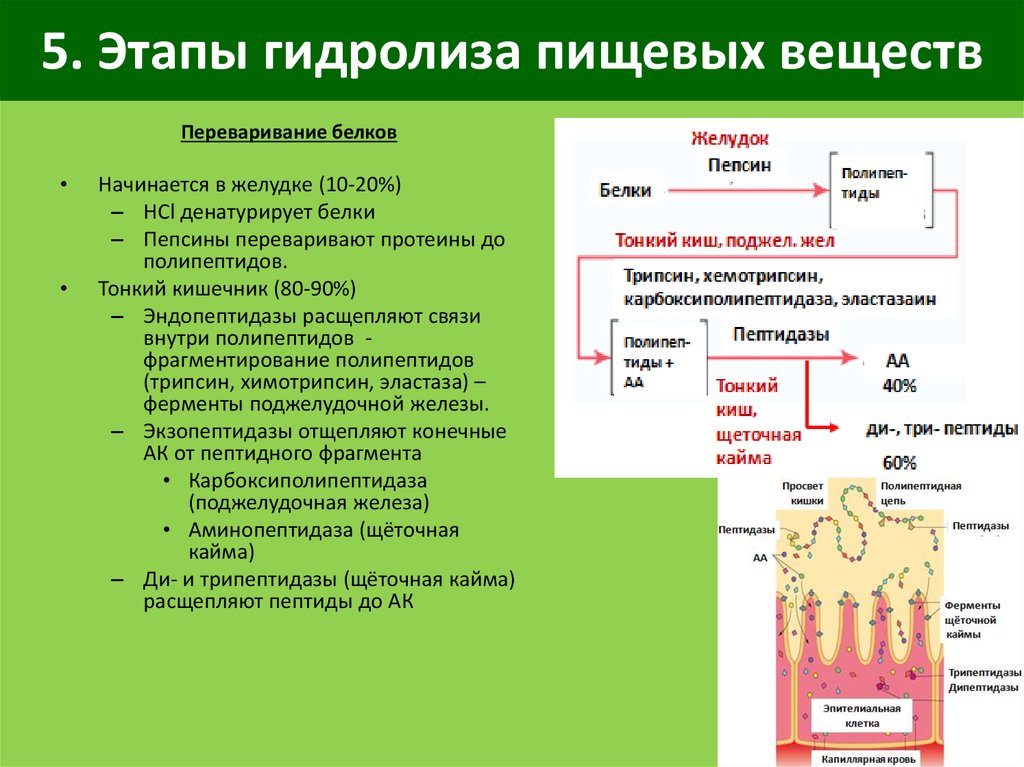
Биологические формы поведения (пищевое, оборонительное, групповое, половое и др.) складываются из многочисленных унитарных (одиночных) действий. Унитарные реакции являются как бы «элементарными частицами» поведения, т.е. это одиночные условные и безусловные рефлекторные акты, составляющие более сложные поведенческие акты. Например, прием корма включает в себя такие унитарные реакции, как выбор пищи (куска или пучка травы), взятие губами, захват зубами, жевание, глотание и так далее.
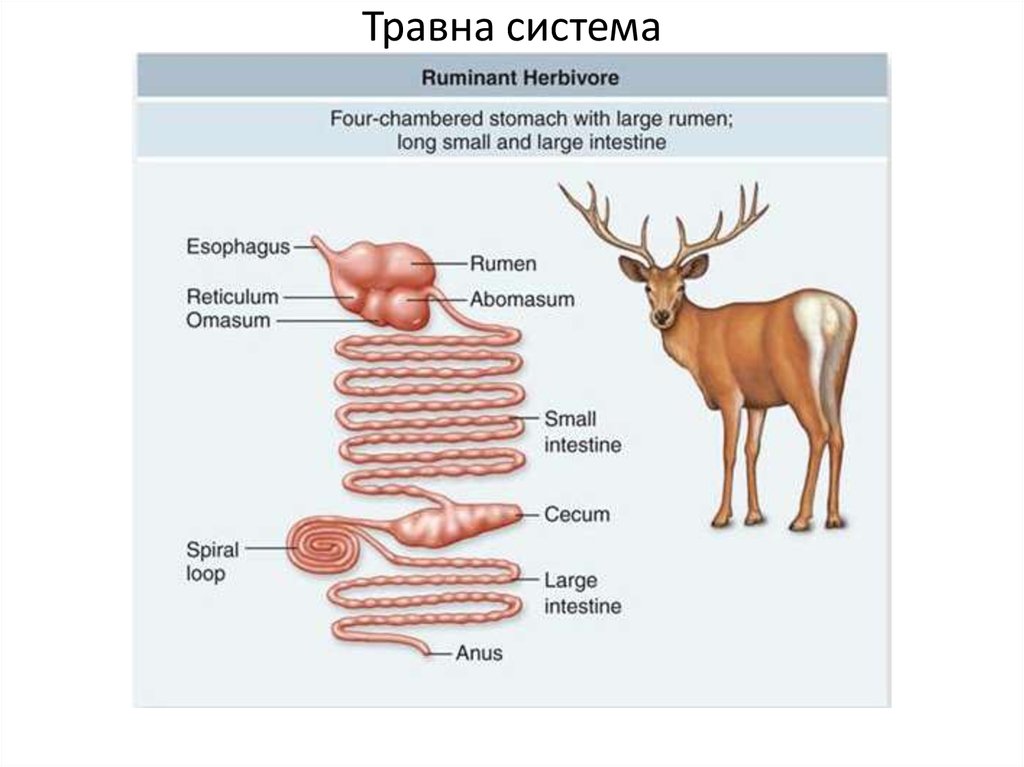
Пищевое поведение складывается из пищедобывательных действий (выбор корма из кормушки, пастьба), собственно приема корма, его обработки, жвачного процесса (у соответствующих видов животных), дефекации.
Одним
из первых безусловных рефлексов,
реализующимся у новорожденных
млекопитающих, является сосательный
рефлекс. Благодаря ему животное уже в
первый час жизни получает молозиво –
незаменимый продукт питания, содержащий
все необходимые питательные вещества,
соли, витамины и иммуноглобулины,
обеспечивающие колостральный иммунитет.
На базе сосательного рефлекс и последующих – слюноотделения, глотания, смыкания пищеводного желоба – у животного вырабатываются индивидуальные условные рефлексы, например на вид сосковой поилки.
Пищевые рефлексы определяют количество и состав поедаемого корма. Это зависит не только от наличия корма, но во многом определяется биологическими потребностями животного, его аппетитом, состоянием обменных процессов. На пастбищах травоядные избирательно поедают разные растения, поэтому не случайно в условиях хороших разнотравных пастбищ у животных нормализуются пищеварительные и обменные процессы, если они были нарушены в стойловый период.
Поведение
животных на пастбище очень разнообразно,
оно обусловлено состоянием травостоя,
наличием источника воды, погодными
факторами, плотностью размещения
животных. У коров периоды пастьбы
чередуются с периодами жвачки и отдыха.
Применение брикетированных и гранулированных кормов удобно с технологической точки зрения. Однако коровы и телята съедают его в меньших количествах, чем обычное сено, затрачивая больше времени; жвачный период сокращается. Обнаруженные изменения в пищевом поведении у крупного рогатого скота при даче гранулированных кормов позволили установить оптимальные соотношения гранулированного и обычного корма в рационе, что повысило усвояемость корма и улучшило работу пищеварительного аппарата.
При
анализе пищевого поведения птиц обращает
на себя внимание выбор объектов для
клевания. Только что вылупившиеся
цыплята способны сразу клевать корм,
причем предпочитая круглые зерна. Куры
видят корм на расстоянии до 6 м, благодаря
хорошему зрению различают величину и
цвет зерен и кормушек.
Куры
видят корм на расстоянии до 6 м, благодаря
хорошему зрению различают величину и
цвет зерен и кормушек.
Таким образом, изучение пищевого поведения служит надежным критерием оценки условий кормления и содержания животных и их корректировки с целью предупреждения заболеваний.
Оборонительное поведение животных чрезвычайно разнообразно. Оно может проявляться по отношению как к живым объектам (животным, людям), так и к неживым предметам, если они сигнализируют о какой – то угрозе для животного или его потомства. Различают две крайние формы оборонительной реакции – пассивная и активная. Пассивная оборонительная реакция включает такие элементы, как бегство, прятанье, оцепенение (неподвижность), а активная – нападение на противника, агрессия.
Групповое
(социальное) поведение.
Внутри группы животных устанавливаются
определенные взаимоотношения, основанные
на законах подчинения и господства
(доминирования). Поэтому любая группа
состоит не просто из отдельных животных,
а представляет собой целостную структуру
– сообщество. В условиях привязного
содержания или в малочисленных группах
групповое поведение не имеет большого
значения. Однако на пастбище, выгуле
или при боксовом содержании социальное
поведение животных проявляется и требует
большего внимания со стороны
зооветспециалистов. Во вновь созданной
большой группе из 20…50 животных сначала
происходит знакомство между особями,
а затем возникают конфликты и соперничество.
В условиях привязного
содержания или в малочисленных группах
групповое поведение не имеет большого
значения. Однако на пастбище, выгуле
или при боксовом содержании социальное
поведение животных проявляется и требует
большего внимания со стороны
зооветспециалистов. Во вновь созданной
большой группе из 20…50 животных сначала
происходит знакомство между особями,
а затем возникают конфликты и соперничество.
Хотя
крупный рогатый скот разного пола и
возраста содержится раздельно, тем не
менее, в каждом сообществе идет
соперничество за лучшее место у кормушки
или поилки, за более удобное место для
отдыха. В результате драк и стычек среди
животных выявляются особи более высокого
ранга (чина) – вожаки и лидеры, и более
низкого ранга – подчиненные. Установившаяся
социальная иерархия сохраняется
достаточно длительное время, и в
сообществе налаживаются мирные отношения,
но каждое животное занимает свою нишу
среди сородичей. Животные низкого ранга
никогда не подойдет первым к кормушке
и не ляжет на самое удобное место, поэтому
в такой группе нет драк и агрессивного
поведения.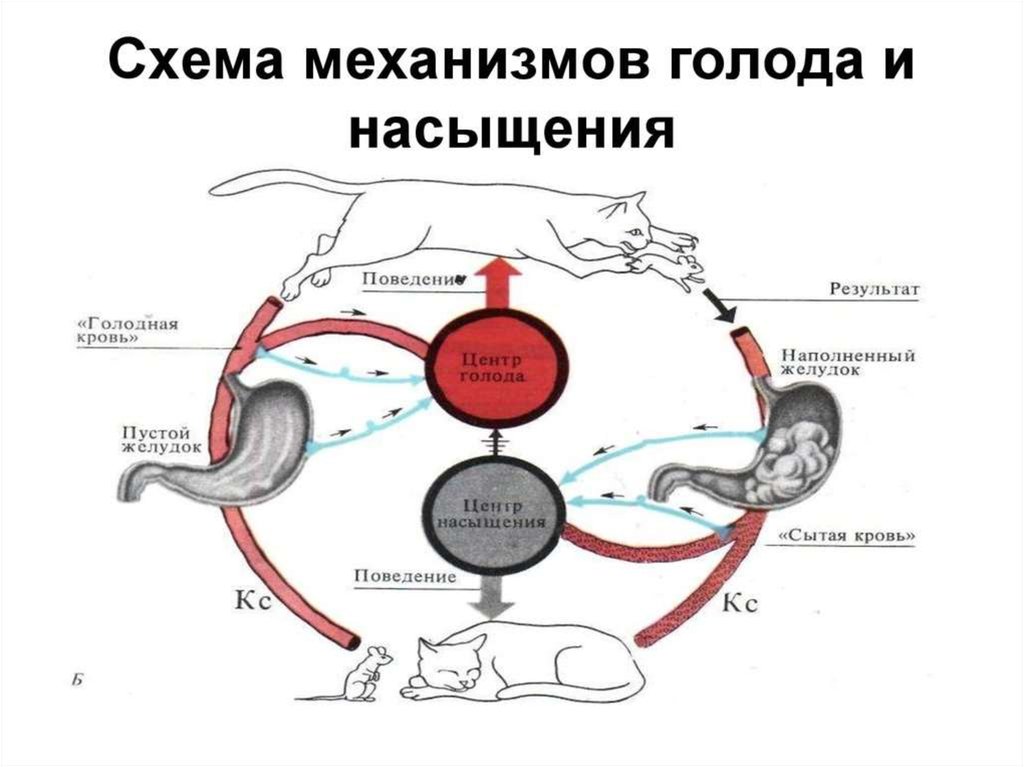 Достаточно угрожающего жеста
со стороны высокорангового животного
– и конфликт будет исчерпан. Кроме
подчиненных в группе находятся и другие
ранги, например «контактные». Это
животные, которые стараются мирно
уживаться со всеми другими, вступают с
ними в дружелюбные отношения (трутся,
облизывают друг друга). Есть в группе и
индифферентные животные – они не борются
за лидерство, но и не боятся высокоранговых
животных.
Достаточно угрожающего жеста
со стороны высокорангового животного
– и конфликт будет исчерпан. Кроме
подчиненных в группе находятся и другие
ранги, например «контактные». Это
животные, которые стараются мирно
уживаться со всеми другими, вступают с
ними в дружелюбные отношения (трутся,
облизывают друг друга). Есть в группе и
индифферентные животные – они не борются
за лидерство, но и не боятся высокоранговых
животных.
Изменения социального ранжирования в группе возможны. Например, если высокоранговое животное заболевает и слабеет, его место займет животное более низкого ранга. Молодые, подрастающие животные стараются спровоцировать доминирующих животных на конфликты и занять их место. Стычки и драки возникают при введении в группу новых животных. Драчливость возрастает и при неблагоприятных условиях содержания: слишком много животных в группе, и они не в состоянии запомнить друг друга, скученность, ограниченный доступ к кормушке или поилке.
Конфликтные
ситуации в группе, поскольку они
разрешаются обычно через драки, приводят
к снижению продуктивности и к повышению
травматизма животных.
Социальное поведение животных в больших группах требует постоянного контроля. Необходимо предупреждать развитие конфликтов между животными, стараться стабилизировать установившуюся социальную иерархию. Драчливых, агрессивных животных целесообразно выбраковать. Хороший способ снизить агрессивность у коров – это удалить или даже опилить острые рога. Очень нежелательно частое изменение состава группы (перегруппировка). Оптимальное число животных в группе зависит от вида животных, например у коров 20…25, у свиней до 20 особей.
Половое
поведение.
Начинает проявляться в период полового
созревания, а до этого животных обоих
полов содержат вместе. Когда уровень
половых гормонов в крови повышается,
начинают складываться новые взаимоотношения. Появляется интерес к противоположному
полу; первые попытки «вспрыгивания»
(маунтнг) являются тренировкой к будущим
половым актам. Животные становятся
легко возбудимыми, драчливыми, у них
снижается аппетит и поедаемость корма.
Такое поведение у телят наступает в
возрасте 6…8 месяцев, у жеребят – 16…18,
у свиней – 5…8 месяцев. С этого времени
самцов и самочек во избежание
преждевременной беременности следует
содержать раздельно.
Появляется интерес к противоположному
полу; первые попытки «вспрыгивания»
(маунтнг) являются тренировкой к будущим
половым актам. Животные становятся
легко возбудимыми, драчливыми, у них
снижается аппетит и поедаемость корма.
Такое поведение у телят наступает в
возрасте 6…8 месяцев, у жеребят – 16…18,
у свиней – 5…8 месяцев. С этого времени
самцов и самочек во избежание
преждевременной беременности следует
содержать раздельно.
Половое
поведение взрослых животных содержащихся
на выпасе или в загоне без привязи
включает в себя поиск и выбор полового
партнера, и собственно половой рефлекс
(эрекцию, обнимательный рефлекс,
совокупление, эякуляция). Половое
влечение проявляется как у самцов, так
и у самок. Соперничество самцов из – за
самок имеет видовые особенности – это
бои (драки), ритуальное поведение,
ухаживание, украшение себя (половой
диморфизм) – изменение в брачный период
окраски, длинной шерсти на отдельных
участках тела или перьев у птиц. В
большинстве случаев самцы, таким образом,
привлекают к себе внимание самки, которой
принадлежит окончательный выбор.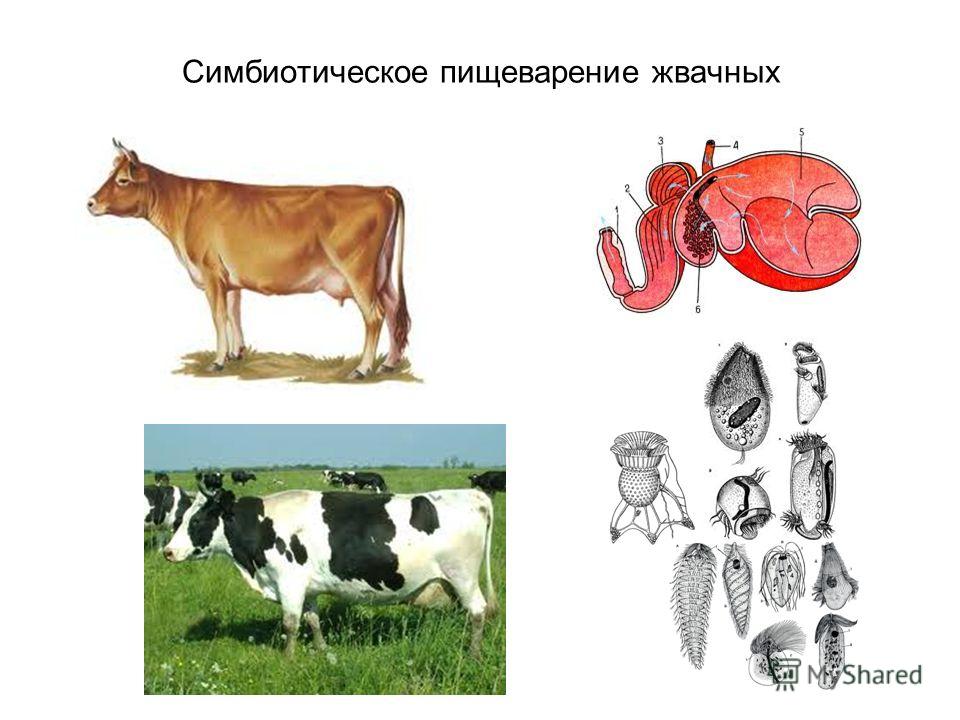
В табуне или стаде самцы отыскивают самок в состоянии половой охоты благодаря органам чувств и, прежде всего – обонянию. В период половой охоты самки и самцы выделяют специфические запаховые половые гормоны – феромоны, которые улавливаются на большом расстоянии (иногда несколько километров). Феромоны стимулируют половое поведение и самцов и самок.
Половые рефлексы у животных направлены на получение полноценного, жизнеспособного потомства. В период размножения половые рефлексы зачастую резко меняют все другие поведенческие реакции: у животных теряется чувство самосохранения, резко снижается поедаемость корма и продуктивность, усиливается агрессивность и неповиновение.
В
условиях искусственного осеменения у
кобыл, коров и свиноматок естественное
половое поведение оказывается
нереализованным, что приводит к снижению
оплодотворяемости. При искусственном
введении спермы в половые органы самки
моторика матки не усиливается, поэтому
спермии не могут достичь рогов матки и
яйцеклетки. Полноценные антиперистальтические
сокращения матки наступают только во
время коитуса, поэтому при искусственном
осеменении используют быков – пробников,
у которых перевязаны семенные канатики.
Такие быки легко отыскивают корову в
состоянии половой охоты и производят
садку, но сразу после этого в половые
пути самки вводят сперму шприцем от
другого, более ценного быка – производителя.
В таком случае оплодотворяемость самок
повышается.
Полноценные антиперистальтические
сокращения матки наступают только во
время коитуса, поэтому при искусственном
осеменении используют быков – пробников,
у которых перевязаны семенные канатики.
Такие быки легко отыскивают корову в
состоянии половой охоты и производят
садку, но сразу после этого в половые
пути самки вводят сперму шприцем от
другого, более ценного быка – производителя.
В таком случае оплодотворяемость самок
повышается.
Материнское поведение. Обеспечивает сохранение, выращивание и обучение потомства. Оно проявляется еще до родов. Беременные животные становятся спокойными, много отдыхают, избегают контакта с другими животными. На пастбищах матки за 2..3 суток до родов часто уходят из стада, прячутся; кобылы готовят сухое ложе в каком – нибудь укромном месте. В условиях стойлового содержания следует размещать самок перед родами в индивидуальных боксах (стойлах).
Во
время родов матери способны самостоятельно
позаботиться о новорожденном, и
ветеринарная помощь нужна лишь в трудных
случаях. Материнские инстинкты включают
в себя облизывание детеныша, массаж его
тела, помощь в поднятии на ноги и
отыскивании вымени, охрану. Материнское
поведение очень хорошо выражено у всех
видов сельскохозяйственных животных
и птиц.
Материнские инстинкты включают
в себя облизывание детеныша, массаж его
тела, помощь в поднятии на ноги и
отыскивании вымени, охрану. Материнское
поведение очень хорошо выражено у всех
видов сельскохозяйственных животных
и птиц.
Быстрое (на 2…3 сутки) отнятие новорожденных и дальнейшее их искусственное кормление и воспитание наносят ущерб здоровью и матери, и детенышей. Ранний отъем телят от матерей технологически обоснован в молочном животноводстве, однако с физиологической точки зрения это нежелательно. Разрабатываются более рациональные технологии содержания телят; например, с использованием коров – кормилиц. Первые 10…11 суток их содержат вместе с матерями, а затем подпускают по 3…4 теленка к лактирующей корове со спокойным нравом и хорошо развитыми материнскими инстинктами.
Комфортное
поведение – это
реакции животных, направленные на
создание для себя благоприятных условий
жизни. К этому типу поведенческих актов
относятся выбор места для отдыха, удобная
поза, купание в воде, валяние в грязи
или песке, поиск укрытий в непогоду,
почесывание кожи и др.
Исследовательское поведение. Одна из важнейших форм поведения, позволяющая изучать и оценивать окружающую среду. Оно включает ориентировочные реакции (ориентация животного на восприятие раздражителя) и манипуляторно – исследовательское поведение. Последнее проявляется в том, что животное оценивает обстановку не просто наблюдая, но и определенным образом воздействуя (облизывает и переворачивает предметы, захватывает зубами).
Изменение пищевого поведения животных в искусственной среде
Человек старается максимально сократить непродуктивные расходы энергии и питательных веществ домашними животными.
Он изолирует животных от контактов с неблагоприятными природно-климатическими факторами, от хищников и возможных источников заболеваний. Животные реагируют на столь радикальное изменение среды обитания прежде всего изменением поведения.
Степень неволи животного может быть различной: от лабораторной клетки до природных резерваций в несколько квадратных километров. В условиях неограниченной свободы животные затрачивают много энергии на локомоции в связи с поиском пищи, воды, уклонением от опасности и пр. Рацион животных в природе может сильно отличаться ото дня ко дню как количественно, так и качественно. На воле животное имеет возможность выбирать кормовые средства.
В условиях неограниченной свободы животные затрачивают много энергии на локомоции в связи с поиском пищи, воды, уклонением от опасности и пр. Рацион животных в природе может сильно отличаться ото дня ко дню как количественно, так и качественно. На воле животное имеет возможность выбирать кормовые средства.
Домашние животные находятся в полной зависимости от человека в части обеспеченности кормами. В неволе животные получают корма, количество и качество которых задает человек. Искусственный рацион домашних животных достаточно однообразен и не меняется на протяжении многих поколений. Такая ситуация приводит к изменению пищевого поведения домашних животных. В пищевых предпочтениях животных вообще и у домашних животных в частности большую роль играет ранний опыт.
На лабораторных животных показано, что уже в раннем постнатальном периоде происходит формирование пищевых предпочтений животных. Если взрослым животным предложить незнакомые кормовые средства более высокого качества вместе с кормами, знакомыми, но с более низкой питательной ценностью, предпочтение отдается последним.
Еще И. П. Павлов подметил эту черту домашних животных. Он разработал методику, которая получила название «сюрпризной пробы». Суть методики заключалась в следующем. Собаку с раннего возраста приучали к однообразной пище — сухарям. Затем в одно из плановых кормлений собаке предлагали вместо привычных сухарей мясо. Мясо собаке было незнакомо, поэтому голодное животное некоторое время пребывало в замешательстве, не зная, как поступить с этой более питательной и естественной для нее пищей. При одновременном предъявлении собаке сухарей и мяса голодное животное без колебаний поедало привычные для него сухари. Требовалось время, чтобы собака попробовала мясо и привыкла к его вкусу.
Пищевую избирательность проявляют все животные без исключения. Однако у домашних животных она выражена резче.
Дикие растительноядные и всеядные животные имеют более широкий спектр пищевых средств из-за постоянного изменения состояния растительного покрова и видового состава кормовых угодий на протяжении года. У хищных видов также имеют место сезонные изменения состава рациона. Из этого следует, что дикие животные плохо переносят однообразие в составе рациона. Однако они так же, как домашние животные, отказываются от незнакомых для них кормов, даже если эти корма имеют высокую питательную ценность.
У хищных видов также имеют место сезонные изменения состава рациона. Из этого следует, что дикие животные плохо переносят однообразие в составе рациона. Однако они так же, как домашние животные, отказываются от незнакомых для них кормов, даже если эти корма имеют высокую питательную ценность.
При содержании диких животных в неестественных для них условиях (ограничение свободы и отсутствие свободного выбора кормовых средств) крайне важно предварительно изучить пищевое поведение и кормовые предпочтения объекта приручения в его естественной среде обитания. Неправильное кормление зачастую становится главной причиной неудачного разведения животных в неволе, например в зоопарках.
В последние годы было предпринято много попыток ввести в аквакультуру новые ценные породы рыб из природных биотопов. Но попытки завершались неудачей главным образом из-за того, что не удалось разработать систему кормления молоди и личинок этих рыб. Например, японцы при одомашнивании камбалы Paralichtys olivaceus столкнулись с тем, что личинка этой рыбы до 10-дневного возраста отказывалась поедать коловраток в присутствии рачка артемии (Artemia sp. ). Но начиная с 25-го дня жизни молодь камбалы отдавала предпочтение уже исключительно артемии. Обнаружилось также, что у личинок раннего возраста ключевым раздражителем пищевой активности выступала плотность коловраток в бассейне. Личинки старшего возраста активизировали пищедобывающее поведение независимо от количества присутствующих в бассейне кормовых объектов.
). Но начиная с 25-го дня жизни молодь камбалы отдавала предпочтение уже исключительно артемии. Обнаружилось также, что у личинок раннего возраста ключевым раздражителем пищевой активности выступала плотность коловраток в бассейне. Личинки старшего возраста активизировали пищедобывающее поведение независимо от количества присутствующих в бассейне кормовых объектов.
Очень непростое пищевое поведение и, соответственно, высокую избирательность пищевых объектов исследователи обнаружили и у других видов. При адаптации к аквариумным условиям серебряного пинагора (Pampus argenteus) выяснилось, что у этой привлекательной для декоративной аквариумистики рыбки активное пищевое поведение и развитие тела происходит лишь в том случае, когда молодь получает живых коловраток в комбинации с микроскопическими водорослями. Собственно микроводоросли не являются жизненно необходимыми кормовыми средствами для молоди этой рыбы. Однако они снижают интенсивность светового потока у поверхности воды и затормаживают фототаксис коловраток. В результате коловратки более равномерно распределяются в толще воды, что делает их более доступными для малоподвижных личинок пинагора.
В результате коловратки более равномерно распределяются в толще воды, что делает их более доступными для малоподвижных личинок пинагора.
Идентичная или даже более высокая питательность искусственных кормов по сравнению с их естественными аналогами не гарантирует нормального роста и развития диких животных в неволе. R. L. Sharma и Е. N. Chakrabarti (1999) сравнивали интенсивность роста личинок карпа (Cyprinus carpio) в искусственных условиях на планктоне и на искусственном корме, в состав которого входили рыбная мука, дрожжи, минерально-витаминные добавки, пшеничная мука и отруби риса. Через 40 дней живая масса личинок, получавших в качестве корма зоопланктонные организмы, была в 3-5 раз выше.
При выращивании осетровых рыб установлено, что как замороженный планктон, так и планктон воздушной сушки по питательности не уступает живому корму. Тем не менее молодь осетра медленнее растет и хуже развивается именно на консервированных кормах. При этом отмечается более высокая смертность личинок осетра.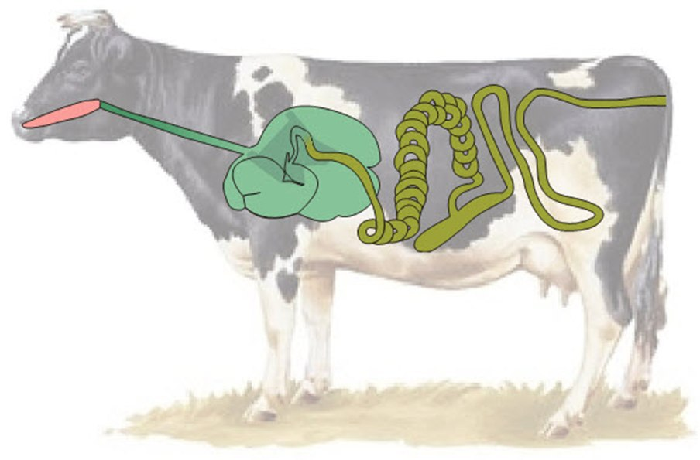 Обнаружилось, что живой корм активизирует у личинок пищеварительные процессы — гидролиз нутриентов, всасывание продуктов гидролиза, усиливает перистальтику кишечника. Предполагается, что важное стимулирующее влияние на процессы роста у личинок могут иметь процессы автолиза, которые развиваются только в живых организмах и отсутствуют в консервированных любым способом кормах.
Обнаружилось, что живой корм активизирует у личинок пищеварительные процессы — гидролиз нутриентов, всасывание продуктов гидролиза, усиливает перистальтику кишечника. Предполагается, что важное стимулирующее влияние на процессы роста у личинок могут иметь процессы автолиза, которые развиваются только в живых организмах и отсутствуют в консервированных любым способом кормах.
В Новой Зеландии ученые пытались одомашнить один из видов палтуса — Colistium nudipinnis. Однако работы с этой ценной рыбой пришлось прекратить из-за того, что не удалось разработать альтернативный искусственный корм для личинок. По аналогичной причине были прекращены работы по одомашниванию и зеленой камбалы Rhombosolea tapirina — очень ценного объекта. Замена естественного дорогого корма на более питательный искусственный рацион сопровождалась 100%-ной гибелью личинок камбалы.
В ряде случаев успех искусственного воспроизводства животных определяется режимом кормления. Результативность выращивания личинок пинагора Cyclopterus lumpus резко повышается, если живой корм задавать рыбе небольшими порциями 2-3 раза в день. Постоянное присутствие в бассейне с личинками зоопланктонных организмов обеспечивает более низкие темпы роста и развития рыбы.
Постоянное присутствие в бассейне с личинками зоопланктонных организмов обеспечивает более низкие темпы роста и развития рыбы.
При содержании животных в неволе необходимо определить и оптимальную физическую форму искусственных кормов. Так, установлено, что цыплята лучше растут, если получают стартовый комбикорм в форме мелких гранул по сравнению с комбикормом того же состава, но в виде влажной мешанки. Потребление гранул сопровождается меньшими энергетическими затратами цыплят при более высокой переваримости белков, жиров и углеводов рациона.
Физическая форма кормовых средств влияет и на поведение жвачных животных. Кормление молочных коров брикетами, приготовленными из смеси цельных растений ячменя и клевера, приводит к изменению поведения животных на фоне коров, получающих цельные растения воздушной сушки или крупную резку растений ячменя и клевера (А. А. Иванов, 1978). У них сокращается время на прием корма, а также сокращается жвачный процесс. В конечном счете, животные больше времени отдыхают. Кормление жвачных животных исключительно гранулированными кормами с размером частиц менее 0,5 см даже при более высокой питательной ценности таких кормов приводит не только к изменению в суточном балансе поведения животных, но и к развитию тяжелых патологических состояний: нарушению руминации и обмена веществ.
Кормление жвачных животных исключительно гранулированными кормами с размером частиц менее 0,5 см даже при более высокой питательной ценности таких кормов приводит не только к изменению в суточном балансе поведения животных, но и к развитию тяжелых патологических состояний: нарушению руминации и обмена веществ.
У некоторых видов диких животных при содержании в искусственных условиях возникают проблемы, с узнаванием кормовых средств. Цыплята страуса голодают при наличии перед ними корма, который они не распознают в качестве такового. Ключевыми раздражителями для страусят являются цвет и форма кормовых средств. Девятидневные птенцы страуса в 10 раз чаще клюют тонкие зеленые полоски с белыми включениями по сравнению с полностью белыми полосками. На аналогичные полоски красного, голубого и желтого цвета птенцы вообще не обращают внимания. Белые полоски на зеленых колбасках ассоциируются у страусят с полосками солей мочевой кислоты, которые видны на каловых массах взрослых страусов. Полоски солей мочевой кислоты в данном случае выступают в качестве знакового раздражителя для капрофагии цыплят, за счет которой они формируют симбиотический микробиологический пейзаж своего желудочно-кишечного тракта после выклева из яйца.
У современных представителей домашних животных произошли заметные изменения поведения вследствие скармливания им больших количеств концентрированных кормов. Быстрое поедание высокопитательного корма делает бессмысленным целый ряд локомоций, присутствующих у диких животных: кормодобывающую активность и связанные с ней миграции, продолжительное жевание, запасание корма, защиту корма от посягательств конкурентов и пр. Место этих поведенческих актов заняли другие стереотипы поведения: отдых, сон, игры, половая активность.
Повышение доли клетчатки за счет включения в рацион сена при содержании жирафа (Giraffa Camelopardalis) в условиях зоопарка привело к тому, что у животных возросло время жвачки и сократилось время видотипичных оральных движений (движений губ, игровых движений языка, облизывания деревьев и других предметов).
Одомашнивание принесло еще одну серьезную проблему — отказ матерей от своих детенышей после родов. Человек был вынужден в таких ситуациях разрабатывать технологии ручного выкармливания новорожденных. К настоящему времени работники зоопарков накопили богатый опыт в этой части. Однако это вовсе не означает, что проблема ручного выкармливания новорожденных решена полностью. Скажем, детеныши некоторых видов млекопитающих отказываются сосать соску. Повторяющиеся насильственные скармливания молока настолько стрессируют новорожденных, что приводят к их гибели. Исследования показали, что стимулом сосательных движений у некоторых видов млекопитающих периода новорожденности выступает запах материнской слюны или запах его собственной слюны. Смазывание соски слюной матери (или собственной слюной новорожденного) стимулирует сосательную активность детеныша. Незнание подобных видоспецифичных ключевых сигналов ставит под угрозу успешное выращивание новорожденных в отсутствие матери.
К настоящему времени работники зоопарков накопили богатый опыт в этой части. Однако это вовсе не означает, что проблема ручного выкармливания новорожденных решена полностью. Скажем, детеныши некоторых видов млекопитающих отказываются сосать соску. Повторяющиеся насильственные скармливания молока настолько стрессируют новорожденных, что приводят к их гибели. Исследования показали, что стимулом сосательных движений у некоторых видов млекопитающих периода новорожденности выступает запах материнской слюны или запах его собственной слюны. Смазывание соски слюной матери (или собственной слюной новорожденного) стимулирует сосательную активность детеныша. Незнание подобных видоспецифичных ключевых сигналов ставит под угрозу успешное выращивание новорожденных в отсутствие матери.
Классическим примером успешного решения проблемы ручного выкармливания диких животных служит опыт североамериканских орнитологов, описанный Б. Кеплером (1978). В связи с уничтожением исторических мест обитания североамериканских журавлей Crus Canadensis и Crus Americana численность этих видов птиц катастрофически сократилась.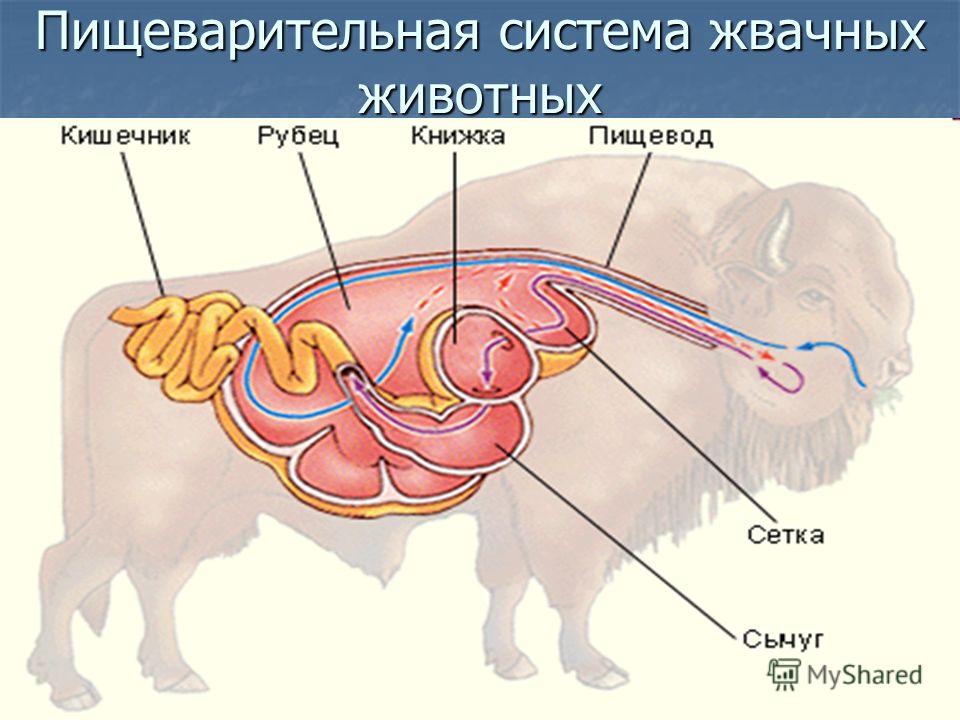 Возникла реальная угроза их исчезновения из природных биотопов. Создание национальных парков и природных резерваций уже не давало положительного результата. Остановить процесс вымирания популяций журавлей могло лишь искусственное воспроизводство птиц. Однако работа по искусственному восстановлению поголовья зашла в тупик из-за того, что после искусственной инкубации яиц и выклева птенцы журавлей не проявляли пищевой активности. Насильственное ручное выкармливание птенцов имело очень низкую результативность: большинство журавлят погибало.
Возникла реальная угроза их исчезновения из природных биотопов. Создание национальных парков и природных резерваций уже не давало положительного результата. Остановить процесс вымирания популяций журавлей могло лишь искусственное воспроизводство птиц. Однако работа по искусственному восстановлению поголовья зашла в тупик из-за того, что после искусственной инкубации яиц и выклева птенцы журавлей не проявляли пищевой активности. Насильственное ручное выкармливание птенцов имело очень низкую результативность: большинство журавлят погибало.
Для птенцов раннего периода были разработаны специальные стартерные комбикорма. Но птенцы не воспринимали комбикорм в качестве кормового средства. Неожиданный результат был получен при выкармливании журавлят вручную смесью стартерного комбикорма и консервированного липкого корма для собак до момента, когда птенцы научались распознавать мешанку в качестве корма. В этих условиях кормовая мешанка прилипала к клюву птенцов, что затрудняло дальнейшее потребление корма, но зато стимулировало у них аппетит и пищевую активность.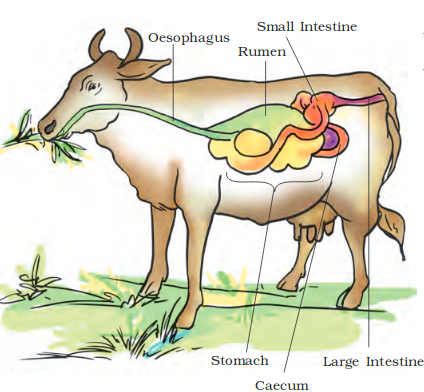 Птенцы начинали склевывать друг у друга прилипший к клюву липкий собачий корм. Только после этого произошло переключение птенцов на искусственный корм в форме мешанки. Однако произошло это событие не сразу. Длительное время внимание журавлят привлекал лишь прилипший собачий корм. При этом птенцы оставались голодными и плохо росли: химический состав собачьего корма не соответствовал потребностям журавлят.
Птенцы начинали склевывать друг у друга прилипший к клюву липкий собачий корм. Только после этого произошло переключение птенцов на искусственный корм в форме мешанки. Однако произошло это событие не сразу. Длительное время внимание журавлят привлекал лишь прилипший собачий корм. При этом птенцы оставались голодными и плохо росли: химический состав собачьего корма не соответствовал потребностям журавлят.
Незадача с искусственным выкармливанием журавлят заставила исследователей обратить более пристальное внимание на пищевое поведение родителей и выклюнувшихся птенцов в их естественной среде обитания. Было установлено, что родители обучают своих птенцов распознавать корм. Для этого родители берут в клюв небольшие кусочки корма и предлагают птенцам. Голодные птенцы обыкновенно выпрашивают у родителей корм, поклевывая их в основание клюва. Через некоторое время взрослые птицы задерживают продемонстрированный кусочек в своем клюве и погружают его в основную часть заготовленного корма у края гнезда. Птенец, продолжая требовать корм, случайно захватывает некоторое количество корма у края гнезда. Через несколько уроков птенец в состоянии голода переключается с клюва родителя на кормовую массу, которую им приносят в гнездо. После этого птенцы продолжают выпрашивать корм через поклевывание родителей в основание клюва. Однако при наличии в гнезде корма они его жадно поедают без процедуры выпрашивания.
Птенец, продолжая требовать корм, случайно захватывает некоторое количество корма у края гнезда. Через несколько уроков птенец в состоянии голода переключается с клюва родителя на кормовую массу, которую им приносят в гнездо. После этого птенцы продолжают выпрашивать корм через поклевывание родителей в основание клюва. Однако при наличии в гнезде корма они его жадно поедают без процедуры выпрашивания.
Наблюдения за поведением родителей и птенцов показали, что в первые часы жизни ключевым раздражителем пищевой активности птенцов служит не сама пища, а родительский клюв. Это наблюдение было использовано и для искусственной стимуляции пищевого поведения журавлят в условиях питомника. С этой целью только что выклюнувшимся журавлятам подставляли деревянную палочку длиной 20 см и 12 мм в диаметре. Такой муляж, особенно красного цвета, провоцировал у птенцов клевательные движения. Если перед птенцами помахать такой палочкой, а затем погрузить ее в корм, то птенцы продолжая клевки, рано или поздно случайно захватывали кусочки корма. Требовалось всего несколько подобных уроков, чтобы переключить внимание журавлят на искусственный корм. Все журавлята обучаются самостоятельно клевать искусственный корм за один час тренировок с использованием муляжа, напоминающего клюв взрослого журавля.
Требовалось всего несколько подобных уроков, чтобы переключить внимание журавлят на искусственный корм. Все журавлята обучаются самостоятельно клевать искусственный корм за один час тренировок с использованием муляжа, напоминающего клюв взрослого журавля.
Дальнейшая модернизация этой методики позволила вообще отказаться от ручного манипулирования искусственным клювом. Жесткая фиксация муляжа в сосуде с мешанкой или его подвешивание над кормом обеспечивает развитие активного пищевого поведения у журавлят в возрасте 1-3 дней. Логика обучения та же, что и при ручном манипулировании искусственным клювом. Муляж распознается птенцами с первых минут жизни вне скорлупы. Птенцы инстинктивно клюют муляж, случайно погружают свой клюв в корм и вскоре начинают распознавать и активно потреблять искусственный комбикорм.
Работы по приручению диких животных свидетельствуют, что большую помощь животным в реализации их адаптивных возможностей оказывает жестко выдерживаемый режим кормления. Приручение диких шимпанзе протекает быстрее и успешнее, когда животных кормят в одно и то же время. Незапланированное внезапное кормление воспринимается обезьянами как дополнительный отрицательный фактор. Для зоопарков режим кормления важен и с другой точки зрения: он сокращает затраты труда обслуживающего персонала, а также представляет возможность посетителям понаблюдать за активными раскованными действиями животных в период кормления. Предсказуемость кормления животных в зоопарке на фоне чувства голода активизирует их поведение, снимает депрессии, что, в конечном счете, повышает их привлекательность для зрителей. Режим кормления помогает животным при групповом содержании оптимизировать их иерархические отношения, понизить агрессивность и травматизм.
Незапланированное внезапное кормление воспринимается обезьянами как дополнительный отрицательный фактор. Для зоопарков режим кормления важен и с другой точки зрения: он сокращает затраты труда обслуживающего персонала, а также представляет возможность посетителям понаблюдать за активными раскованными действиями животных в период кормления. Предсказуемость кормления животных в зоопарке на фоне чувства голода активизирует их поведение, снимает депрессии, что, в конечном счете, повышает их привлекательность для зрителей. Режим кормления помогает животным при групповом содержании оптимизировать их иерархические отношения, понизить агрессивность и травматизм.
Режим кормления имеет не меньшее значение и для домашних животных. В молочном скотоводстве задержка с раздачей кормов приводит к снижению надоев молока. В яичном птицеводстве нарушение графика кормораздачи задерживает яйцекладку. При групповом содержании свиней бессистемная раздача кормов стимулирует агрессивность у животных, следствием чего выступает травматизм, снижение среднесуточных привесов, перерасход кормов.
С другой стороны, имеется статистика наблюдений, которая указывает на то, что освобождение животных от пищедобывающей активности увеличивает у них долю свободного времени. Этот результат не всегда рассматривается как положительное явление. Зачастую свободное время животные занимают драками или избыточной неподвижностью (сон и отдых). Это отрицательно сказывается на общем состоянии животных. Если в условиях ограниченной свободы животные имеют много свободного времени и демонстрируют патологические формы поведения, агрессию, их свободное время, возможно, следует ограничить. С этой целью разрабатывались специальные устройства, которые вынуждали животных совершать дополнительные движения при кормлении.
Для свиней в свое время было предложено изобретение, получившее название «эдинбургского футбола». Гранулированный комбикорм заправляли в металлический круглый резервуар (мяч) с небольшими отверстиями. Для того чтобы из резервуара высыпался корм, его надо перекатывать с места на место. Данный метод кормораздачи заметно увеличивал продолжительность потребления корма свиньями, активизировал их двигательную активность, сокращал период сна и отдыха. Ограничение свободного времени привело и к сокращению агрессивных действий животных.
Данный метод кормораздачи заметно увеличивал продолжительность потребления корма свиньями, активизировал их двигательную активность, сокращал период сна и отдыха. Ограничение свободного времени привело и к сокращению агрессивных действий животных.
Похожий кормораздатчик применяется в коневодстве для увеличения продолжительности времени, затрачиваемого лошадьми на пищевое поведение.
Для стимуляции мышечной и психической активности в процессе кормления применяются и другие приемы. Так, классическая скиннеровская камера является не только инструментом научения и выработки условных рефлексов. При решении поставленной задачи животные в камере вынуждены активизировать поисково-исследовательское поведение. Правильное решение подкрепляется дачей порции корма.
Для профилактики гипокинезии и ожирения домашних собак полезно кормить из механических кормушек, которые дозирование выдают сухой гранулированный корм только после нажатия на специальный рычаг или заслонку. О. Price (2002) сообщает, что данный прием кормления существенно изменяет характер поведения собаки. Собаки вынуждены больше двигаться, они меньше надоедают своим хозяевам, поскольку их свободное время сокращается на фоне более полного удовлетворения потребности в движениях.
Собаки вынуждены больше двигаться, они меньше надоедают своим хозяевам, поскольку их свободное время сокращается на фоне более полного удовлетворения потребности в движениях.
Искусственная стимуляция пищедобывающей активности в ограниченном пространстве особенно полезна в случае, когда животных невозможно заставить выполнять физические нагрузки. Особые трудности возникают при разведении в неволе хищных животных (пауки, ящерицы, птицы, кошки). Неволя радикально изменяет их образ жизни и ломает алгоритмы охотничьего поведения. В природе охотничье поведение, кроме голода, провоцируется и особыми ключевыми раздражителями, которые в неволе отсутствуют или же подавляются раздражителями другого рода.
Иногда существует необходимость стимулировать активные действия для демонстрации животных зрителям в активном состоянии, в движении (зоопарки, аквапарки). Например, американский черный медведь (Ursus americanus) в зоопарке всегда собирает много зрителей, поскольку имеет привлекательную внешность и может продемонстрировать интересные локомоции. Однако в условиях ограниченного пространства он малоактивен, много спит. Ученые разработали способ, который повышает активность медведя, улучшает его общее состояние, а потому собирает массы зрителей. Его кормят особым способом: пищу прячут в норы, заваливают бревнами или маскируют камнями. Животные вынуждены много времени уделять поиску пищи, ее извлечению из-под завалов. Данный прием кормления пролонгирует время поисковой активности медведей. Все это происходит на глазах у восторженных зрителей.
Однако в условиях ограниченного пространства он малоактивен, много спит. Ученые разработали способ, который повышает активность медведя, улучшает его общее состояние, а потому собирает массы зрителей. Его кормят особым способом: пищу прячут в норы, заваливают бревнами или маскируют камнями. Животные вынуждены много времени уделять поиску пищи, ее извлечению из-под завалов. Данный прием кормления пролонгирует время поисковой активности медведей. Все это происходит на глазах у восторженных зрителей.
G. S. Shepherdson et al. (1993) провели наблюдения за поведением кошки-рыболова (Felis viverrina). В эксперименте животные получали корм (рыбу) из кормушки либо были вынуждены самостоятельно ловить рыбу в пруду. В последнем случае время, затрачиваемое на пищедобывающую активность, возрастало до 40% в бюджете дневного поведения кошек. Время на сон, наоборот, сократилось с 67% до 21%.
Если макакам в подстилку подмешивать зерна злаков, то животные много времени занимаются переборкой подстилки. При этом частота агрессивных действий животных сокращается в 5 раз.
При этом частота агрессивных действий животных сокращается в 5 раз.
На воле мелкие кошки, например леопардовая кошка (Felis bengalensis), питаются мелкими животными. Суммарное время охоты за день у них достигает значительной величины. В условиях зоопарков организовать охоту животным не представляется возможным. Однако и в условиях ограниченной свободы можно заставить этих кошек двигаться в 3 раза больше по времени. С этой целью суточный рацион животных делят на 4 части и в соответствии с принятым графиком кормления корм прячут в разных участках вольеры под кучами хвороста, листвы или мусора. Такая кормораздача резко повышает поисковую активность кошек. Животные больше двигаются, сокращают время на стереотипические патологические движения (pacing activity). В конечном счете это положительно отражается на внешнем виде животных и привлекает внимание зрительской аудитории. Кошки пребывают в активном состоянии на протяжении всего дня, а не только перед раздачей корма. При этом на 50% сокращается время, которое кошки обыкновенно затрачивают на бесцельное хождение взад-вперед (9% против 18% времени всех активных действий). Действия животных приближены к охотничьему поведению. Подобные изменения в поведении животных этологи называют натурализацией поведения в неволе.
Действия животных приближены к охотничьему поведению. Подобные изменения в поведении животных этологи называют натурализацией поведения в неволе.
Кормление животных, ведущих групповой образ жизни, имеет свои специфические проблемы. При раздаче кормов первыми питаются доминирующие особи. Субдоминанты получают то, что не съедают высокоранжированные особи группы. В результате доминанты переедают и жиреют, а субдоминанты остаются голодными. Частично проблема решается за счет применения кормушек, поделенных на сектора.
M.-F. Bouisson (1970) продемонстрировал эффективность этого приема на крупном рогатом скоте при парном кормлении животных, занимающих в стаде разное иерархическое положение.
Таким образом, очевидно, что пищевое поведение животных в неволе существенно изменяется. В зависимости от решаемых человеком задач технологией кормления можно как ограничивать поведенческую активность животных, так и стимулировать ее.
Пищевое поведение и связанная с ним деятельность у жвачных животных
 «>
«> Arnold, G.W. and Dudzinski, M.L. (1978). Этология домашних животных, живущих на свободном выгуле . (Амстердам, Оксфорд, Нью-Йорк: Elsevier)
Google Scholar
-
Бэйл, К.А. и Майер, Дж. (1970). Гипоталамические центры: обратные связи и рецепторы в краткосрочном контроле потребления корма. В AT Phillipson (ред.) Физиология пищеварения и метаболизма жвачных животных . Материалы Третьего международного симпозиума, Кембридж, Англия, стр. 254–263. (Ньюкасл-апон-Тайн: Oriel Press)
Google Scholar
-
Балч, CC (1952). Факторы, влияющие на использование корма дойными коровами. 6. Скорость сокращения ретикулума. руб. Дж. Нутр ., 6 , 366–375
CrossRef пабмед КАС Google Scholar
-
Балч, CC (1958). Наблюдения за актом приема пищи у крупного рогатого скота.
 руб. Дж. Нутр ., 12 , 330–345
руб. Дж. Нутр ., 12 , 330–345 CrossRef пабмед КАС Google Scholar
-
Балч, К.С. и Кэмплинг, Р.К. (1962). Регуляция произвольного приема пищи у жвачных животных. Нутр. Абстр. ., 32 , 669–686
PubMed КАС Google Scholar
-
Балч, CC (1971). Предложение использовать время, затрачиваемое на жевание, в качестве показателя степени, в которой рационы для жвачных животных обладают физическим свойством волокнистости, характерным для грубых кормов. руб. Дж. Нутр ., 26 , 383–392
CrossRef пабмед КАС Google Scholar
-
Баумгардт, Б. Р. (1970). Регулирование потребления корма и энергетического баланса. В AT Phillipson (ред.) Физиология пищеварения и метаболизма жвачных животных .
 Труды Третьего международного симпозиума, Кембридж, Англия, стр. 235–253. (Ньюкасл-апон-Тайн: Oriel Press)
Труды Третьего международного симпозиума, Кембридж, Англия, стр. 235–253. (Ньюкасл-апон-Тайн: Oriel Press) Google Scholar
-
Байнс, Дж. А. и Дэйви, А. В. Ф. (1970). Произвольное потребление, пищеварение, скорость прохождения, количество материала в пищеварительном тракте и поведение у коров, получающих полноценные рационы, содержащие солому и концентраты в различных пропорциях. руб. Дж. Нутр ., 24(4), 1013–1018
Перекрёстная ссылка пабмед КАС Google Scholar
-
Брамби, П.Дж. (1959). Пастбищное поведение молочного скота в зависимости от молочной продуктивности, живой массы и потребления пастбищ. Н.З.Дж. Агр. Рез ., 2 , 797–807
CrossRef Google Scholar
-
Берт, AWA (1957). Влияние изменений в потреблении питательных веществ на надои и состав молока.
 II. Факторы, влияющие на скорость поедания грубых кормов и реакцию на увеличение количества скармливаемых концентратов. Дж. Молочная. Рез ., 24 , 296–315
II. Факторы, влияющие на скорость поедания грубых кормов и реакцию на увеличение количества скармливаемых концентратов. Дж. Молочная. Рез ., 24 , 296–315 CrossRef КАС Google Scholar
-
Кэмплинг, RC (1966). Влияние концентратов на скорость исчезновения пищеварения из пищеварительного тракта коров, получавших сено. J. Dairy Res ., 33 , 13–23
CrossRef Google Scholar
-
Кэмплинг, RC (1970). Физическая регуляция произвольного приема пищи. В А.Т. Филлипсон (ред.) Физиология пищеварения и обмена веществ у жвачных . Труды Третьего международного симпозиума, Кембридж, Англия, стр. 226–234. (Ньюкасл-апон-Тайн: Oriel Press)
Google Scholar
-
Чакон, Э. и Стоббс, Т. Х. (1976). Влияние прогрессирующей дефолиации травяного покрова на пищевое поведение крупного рогатого скота.
 австр. Дж. Агри. Рез ., 27 , 709–727
австр. Дж. Агри. Рез ., 27 , 709–727 CrossRef Google Scholar
-
Чейз Л.Э., Вангснесс П.Дж. и Баумгардт Б.Р. (1976). Кормовое поведение бычков, получавших полный смешанный рацион. J. Dairy Sci ., 59 , 1923–1928
CrossRef пабмед КАС Google Scholar
-
Коуэн, А. Т. (1975). Время выпаса и характер выпаса фрезийских коров на тропических травяно-бобовых пастбищах. австр. Дж. Эксп. Агри. Аним. Узб ., 15 , 32–37
Перекрёстная ссылка Google Scholar
-
Демаркильи, К. (1972). Digestibilité, valeur nutritive et ingestibilité des betteraves de différentes teneurs en matière sèche. Энн. Зоотех ., 21 , 415–428
CrossRef Google Scholar
 «>
«> Демаркильи, К. и Далфи, Дж. П. (1977). Влияние силосования на потребление корма и продуктивность животных. В: Материалы Международного совещания по животноводству на пастбищах умеренного пояса , Дублин: Ирландское общество пастбищ.
Google Scholar
-
Десуисен, А., Ванбель, М. и Фокант, М. (1978). Влияние длины измельчения силоса на произвольное потребление и поведение овец при жевании. Брит. Грассл. Soc ., 33 , 107–115
CrossRef Google Scholar
-
Дакворт, Дж. Э. и Ширлоу, Д. В. (1958). Изучение факторов, влияющих на потребление корма и пищевое поведение крупного рогатого скота. Аним. Поведение ., 6 , 147–154
CrossRef Google Scholar
-
Далфи, Дж. П. (1971). Влияние дю poids vif du niveau d’ingestion sur Ie comportement alimentaire et mérycique du mouton.
 Энн. Зоотех ., 20 , 477–486
Энн. Зоотех ., 20 , 477–486 CrossRef Google Scholar
-
Далфи, Дж. П. и Демаркиль, К. (1973). Влияние Ia machine de récolte et de Ia Finesse de hachage sur Ia valeur alimentaire des silages. Энн. Зоотех ., 22 , 199–217
CrossRef Google Scholar
-
Далфи, Дж. П. и Демаркилли, К. (1974). Etude du comportement alimentaire et mérycique de moutons recevant des fourrages verts hachés. Энн. Зоотех ., 23 , 193–212
Google Scholar
-
Далфи, Дж. П., Беше, Г. и Томсон, Э. (1975). Влияние структуры, телосложения и качества сохранения силосных злаков на их пищеварение. Энн. Зоотех ., 24(1), 81–94
CrossRef Google Scholar
 «>
«> Далфи, Дж. П. и Беше, Г. (1976). Влияние дю Stade de vegétation et de l’espèce vegétale sur Ie comportement alimentaire et mérycique de moutons recevant des fourrages verts hachés. Энн. Зоотех ., 25 , 505–519
CrossRef Google Scholar
-
Фонтено, Дж. П. и Блазер, Р. Э. (1965). Симпозиум по факторам, влияющим на добровольное потребление травы жвачными животными: отбор и потребление пастбищными животными. Дж. Аним. Науки ., 24 , 1202–1208
Google Scholar
-
Форбс, Дж. М., Райт, Дж. А. и Баннистер, А. (1972). Заметка о скорости питания овец. Аним. Изд ., 15 , 211–214
CrossRef Google Scholar
-
Freer, M., Campling, RC и Balch, C.C. (1962). Факторы, влияющие на произвольное потребление корма коровами.
 4. Поведение и ретикулярная подвижность коров, получающих рационы из сена, овсяной соломы и овсяной соломы с мочевиной. руб. Дж. Нутр ., 16 , 279–295
4. Поведение и ретикулярная подвижность коров, получающих рационы из сена, овсяной соломы и овсяной соломы с мочевиной. руб. Дж. Нутр ., 16 , 279–295 CrossRef пабмед КАС Google Scholar
-
Фрир, М. и Кэмплинг, Р. К. (1965). Факторы, влияющие на произвольное потребление корма коровами. 7. Поведение и ретикулярная подвижность коров, получавших рационы, сухую траву, концентраты и молотое гранулированное сено. руб. Дж. Нутр ., 19 , 195–207
CrossRef пабмед КАС Google Scholar
-
Жоффруа, Ф. (1974). Сравнительный этюд пищевого и меристичного отношения к двум мелким жвачным животным: шевре и мутону. Энн. Зоотех ., 23(1), 63–73
CrossRef Google Scholar
-
Гилл, Дж., Кэмплинг, Р.К. и Вестгарт, Д.
 Р. (1966). Исследование жевания во время еды у коровы. руб. Дж. Нутр ., 20 , 13–23
Р. (1966). Исследование жевания во время еды у коровы. руб. Дж. Нутр ., 20 , 13–23 CrossRef пабмед КАС Google Scholar
-
Гордон, Дж. Г. (1958). Акт размышления. Дж. Сельское хозяйство. Sci ., 50(1), 34–42
Перекрёстная ссылка Google Scholar
-
Гренет, Э. (1970). Taille и др структура растительных частиц au niveau du feuillet et des fécès chez Ies bovins. Энн. биол. Аним. биох. Биофиз ., 10 ,(4), 643–657
CrossRef Google Scholar
-
Hafez, ESE, Schein, MW, Ewbank, R. (1969). Поведение крупного рогатого скота. В ESE Hafez (ред.) Поведение домашних животных , стр. 235–295. (Лондон: Бальер, Тиндаль и Кассель)
Google Scholar
 «>
«> Хэнкок, Дж. (1953). Пастбищное поведение крупного рогатого скота. Аним. Порода. Абстр. ., 21(1), 1–13
Google Scholar
-
Хэнкок, Дж. (1954). Исследования пастбищного поведения в связи с управлением пастбищами. I. Различия в привычках выпаса молочного скота. Дж. Агри. Sci ., 44 , 420–433
CrossRef Google Scholar
-
Харумото, Т. (1974). Вариабельность жевательного поведения у овец, получавших постоянный рацион. Бык. Фейри. Агри. Shimane Univ ., 7 , 55–79. Абстр. в: биол. Абслр ., 59(10), 5750
Google Scholar
-
Харумото Т. и Като М. (1975). Вариабельность жевательного поведения у коров, получавших постоянное количество сена. Бык. Фейри. Агри. Shimane Univ ., 8 , 15–21. Абстр.
 в: биол. Abstr ., 61(2), 618
в: биол. Abstr ., 61(2), 618 Google Scholar
-
Жарридж Р., Демаркильи К., Журне М. и Беранже К. (1973). Питательная ценность переработанных обезвоженных кормов с особым упором на влияние физической формы и размера частиц. В К. Л. Скидмор и Э. Пламридж (ред.) Труды первого Международного конгресса по сушке зеленых культур , стр. 99–118. (Кембридж: Линтон)
Google Scholar
-
Джарридж, Р., Моранд-Фер, П. и Ходен, А. (1978). Consommation d’aliments et d’eau. В: INRA, Alimentation des Ruminants , стр. 177–206. Эд. Публикации INRA (Route de St-Cyr), 78000 Versailles
Google Scholar
-
Журне, М. и Ремонд, Б. (1976). Физиологические факторы, влияющие на произвольное потребление корма коровами: обзор. Прямой эфир. Произв. Sci .
 , 3 , 129–146
, 3 , 129–146 CrossRef Google Scholar
-
Кербаа, А. (1963). Etude de quelques facteurs de variation du comportement alimentaire et merycique des ruminants. Тезе де Докторат ö’Университет . (Университет Клермон-Ферран)
Google Scholar
-
Лофгрин, Г. П., Мейер, Дж. Х. и Халл, Дж. Л. (1957). Модели поведения овец и крупного рогатого скота, которых кормят пастбищем или силосом. Дж. Аним. ., 15 , 773–780
Google Scholar
-
Мец, JHM (1975). Временные закономерности кормления и жвачки домашнего скота. Мед. Landbouwhogesch. Вагенинген , 45 , 75–12
Google Scholar
-
Оуэн Г.Л., Марц Ф.А., Кэмпбелл Дж.
 Р., Матчи А.Г. и Хильдебранд Э.С. (1976). Связь питания и связанных с ним моделей поведения крупного рогатого скота в условиях содержания с кормовыми видами и температурой окружающей среды. Дж. Аним. Науки ., 42 ,1534–1540
Р., Матчи А.Г. и Хильдебранд Э.С. (1976). Связь питания и связанных с ним моделей поведения крупного рогатого скота в условиях содержания с кормовыми видами и температурой окружающей среды. Дж. Аним. Науки ., 42 ,1534–1540 Google Scholar
-
Пейн, WJA (1966). Питание жвачных в тропиках. Нутр. Абстр. ., 36 , 653–670
PubMed КАС Google Scholar
-
Пети, М. (1972). Emploi du temps des troupeaux de vaches mères et leurs veaux sur Ies pâturages d’Altitude de l’Aubrac. Энн. Zooteeh ., 21(1), 5–27
CrossRef Google Scholar
-
Пятковски Б. и Нагель С. (1975). Ergebnisse über die Kau und Wiederkauaktivität von Kühen bei Rationen mit Getreidestroh verschiedener physikalischer Form und nach chemischer Behandlung mit Natronlange.
 Арех. Тирернер ., 25 , 575–582
Арех. Тирернер ., 25 , 575–582 CrossRef КАС Google Scholar
-
Пятковски Б., Нагель С. и Бергнер Э. (1977). Das Wiederkauverhalten von Kühen bei Unterschiedlicher Trockensubstanzaufnahme und verschiedenen physikalische Form von Grasheu. Арех. Tiererhähr ., 27 , 563–569
CrossRef Google Scholar
-
Патнэм, П. А. и Дэвис, Р. Э. (1963). Влияние рациона на схему кормления бычков на сухостойных стоянках. Дж. Аним. ., 22 , 437–443
Google Scholar
-
Патнэм, П.А., Леманн, Р. и Дэвис, Р.Э. (1964). Норма потребления корма и живая масса мясного скота. Дж. Аним. ., 23 , 425–429
Google Scholar
-
Патнэм П.
 А., Леманн Р. и Любер В. (1968). Суточная норма потребления корма бычками на сухостойном участке. Дж. Аним. ., 27 , 1494–1496
А., Леманн Р. и Любер В. (1968). Суточная норма потребления корма бычками на сухостойном участке. Дж. Аним. ., 27 , 1494–1496 Google Scholar
-
Ray, D.E. и Roubicek, C.B. (1971). Поведение откормочного скота в течение двух сезонов. Дж. Аним. Sci ., 33(1), 72–76
PubMed КАС Google Scholar
-
Ремонд, Б. (1969). Влияние d’un apport croissant d’orge sur Ie comportement alimentaire de la vache laitière et la пищеварение. Энн. Zooteeh ., 18(1), 55–64
CrossRef Google Scholar
-
Ракебуш, Ю. и Бост, Дж. (1963). Etude comparée de la motricité du reticulum et du comportement alimentaire chez Ies ovins et Ies caprins en stabulation. Rev. Méd. Вет ., 114 , 184–196
Google Scholar
 «>
«> Ruckebusch, Y. и Marquet, JP (1963). Recherches sur Ie comportement alimentaire chez Ies ruminants. Ред. мед. Вет ., 114 , 833–856
Google Scholar
-
Ruckebusch, Y., Fargeas, J. and Dumas, J.P. (1970). Recherches sur Ie comportement alimentaire des ruminants. IX. La Mérycique жевания. Rev. Méd. Вет ., 33(4), 345–357
Google Scholar
-
Рукебуш, Ю. и Буэно, Л. (1972). Проанализируйте количественные отношения между активами пищевой промышленности и бдительностью по сравнению со стационарным скотоводством. Энн. Реч. Ветер ., 3(3), 399–420
Google Scholar
-
Ракебуш, Ю. (1975). Моторика желудка жвачных связана с состояниями сна. В IW McDonald and ACI Warner (eds.) Пищеварение и метаболизм у жвачных животных .
 Материалы IV Международного симпозиума по физиологии жвачных животных, стр. 77–90. (Армидейл, Новый Южный Уэльс: издательство Университета Новой Англии)
Материалы IV Международного симпозиума по физиологии жвачных животных, стр. 77–90. (Армидейл, Новый Южный Уэльс: издательство Университета Новой Англии) Google Scholar
-
Ракебуш, Ю. и Буэно, Л. (1978). Анализ пищевого поведения и активности крупного рогатого скота в полевых условиях. Заяв. Аним. Этол ., 4 , 301–313
CrossRef Google Scholar
-
Скоури, М. (1966). Valeur nutritive de la ration et comportement alimentaire du ruminant. Эти Доктор-Инженер. Париж
Google Scholar
-
Судзуки С., Шинде Ю. и Хидари Х. (1970). Влияние изменения ежедневного времени доступа к сену на скорость поедания и потребление корма дойными коровами. яп. Дж. Зоотех. ., 41(8), 423–429
Google Scholar
 «>
«> Судзуки С., Шинде Ю. и Хидари Х. (1973). Влияние раздельного кормления грубыми кормами на скорость поедания молочных коров во время еды. яп. Дж . Зоотех. наука ., 44(3), 181–187
Google Scholar
-
Териес, М., Беше, Г. и Моленат, Г. (1979). Пищевое поведение овец на пастбище. 30-я ежегодная встреча Европейской ассоциации животноводства, Харрогейт, Англия
Google Scholar
-
Томас С. и Кэмплинг Р. К. (1977). Сравнение некоторых факторов, влияющих на усвояемость у овец и коров. Дж.Бр. Грассл. Соэ ., 32 , 33–41
CrossRef Google Scholar
-
Верите Р. и Журне М. (1970). Влияние де-ла-teneur en eau et de la déshydratation de l’herbe sur sa valeur alimentaire pour Ies vaches laitières. Энн. Zooteeh ., 19 , 255–268
CrossRef Google Scholar
 «>
«> Верите, Р. и Журне, М. (1973). Использование Quantités élevées de betteraves par Ies vaches laitières: etude de l’ingestion, de la пищеварение и des eflets sur la production. Энн. Зоотех ., 22(2), 219–235
CrossRef Google Scholar
-
Voskuil, G.C.J. и Metz, JHM (1973). Влияние измельченного сена на потребление корма, скорость поедания и жвачку молочных коров. Нет. Дж. Агрик. ., 21 , 256–262
Google Scholar
-
Уэлч, Дж. Г. и Смит, А. М. (1969). Влияние разного количества корма на жвачку. Дж. Аним. ., 28 , 827–830
Google Scholar
-
Уилсон Р.К. и Флинн А.В. (1974). Наблюдения за пищевым поведением мясного скота, которого кормили индивидуально, предлагали травяной силос вволю. Айриш Дж. Агрик.
 Рез ., 13(3), 347–349
Рез ., 13(3), 347–349Google Scholar
-
Уилсон Р.К. и Флинн А.В. (1976). Пищевое поведение бычков, которым вволю предлагали травяной силос в кормушках с добавлением ячменя и без него. Проц. Нутр. Soc ., 35 (1), 15A-16 A
CrossRef пабмед КАС Google Scholar
-
Ксанде, А. (1978). Valeur alimentaire des pailles de céreales chez Ie mouton. II. Влияние l’espèce de Ia variété et du séjour на Ie sol avant ramassage, sur Ia valeur alimentaire des pailles de céréales. Энн. Зоотех ., 27(4), 601–616
CrossRef Google Scholar
Скачать ссылки
Кормление и жевательное поведение жвачных животных.
- Поиск ресурса VetMed
- Умный поиск
Доступ к более чем 2,4 миллионам рефератов, более 180 000 полнотекстовых документов, а также новостных статей, глав в книгах и описаниям болезней
Введите ключевое слово или фразу
Расширенный библиографический поиск
Интеллектуальный поиск VetMed Resource основан на часто исследуемых темах и ваших собственных запросах.
Запросить поиск
- Агрессия у кошек
- Аллергические реакции у собак
- Анестезия рыб
- Антрозоология
- Летучие мыши и возникающие/зоонозные вирусы
- Поведенческие проблемы попугаев
- Измерение артериального давления (кошки и собаки)
- Здоровье костей кур-несушек
- Туберкулез крупного рогатого скота и барсуков
- Туберкулез крупного рогатого скота и вакцинация
- Заворот-дилатация желудка у собак
- CPR/CPCR собак или кошек
- Вакцины E. coli O157 для крупного рогатого скота
- Остеоартрит лошадей
- Клевание перьев
- Кошачья астма
- Пищевая аллергия/непереносимость у собак и кошек
- Переломы жеребят
- Газопузырьковая болезнь рыб
- Haemonchus contortus – разработка вакцины
- Травмы головы у собак и кошек
- Вирус гепатита Е у свиней
- Неадекватное выделение у кошек
- Герпесвирус кои
- Лечение почечной недостаточности у кошек
- Порок митрального клапана у собак
- MRSA у свиней
- Некротический энтерит и пробиотики/конкурентное исключение
- Новообразования предстательной железы
- Питание и болезни сердца у собак и кошек
- Лечение зубов домашних животных и заболевания полости рта
- Профилактика/контроль РРСС
- Рецидивирующая обструкция дыхательных путей у лошади
- Скрининг собак на наследственные заболевания
- Анестезия мелких млекопитающих
- Стереотипы лошадей
- Оглушение и благополучие домашней птицы
- Свиной грипп и здоровье человека
- Зубные болезни кроликов
- Тотальная внутривенная анестезия (TIVA) собак и кошек
- Мочекаменная болезнь — лечение собак и кошек
- Прививки кошек
- Ветеринарная профессиональная этика
- Лечение ран у лошадей
Подпишитесь, чтобы получать наш электронный информационный бюллетень по ветеринарии и зоотехнике, оповещения о бронировании и предложения прямо на ваш почтовый ящик.
Глава книги CABI
Описание
Одомашнивание позвоночных основано на понимании потребностей животных в их естественной среде, и успех этого одомашнивания на протяжении всей истории человечества во многом зависит от знания их пищевого поведения. Цель этого тома — предоставить продвинутым студентам и исследователям обзор современных знаний о питании домашних млекопитающих и птиц. Первая ф…
Подробнее об этой книге
Глава 14 (Страница №: 241)
Кормление и жевательное поведение жвачных животных.
В этой главе рассматриваются особенности пищевого поведения и жевания у жвачных животных и их значение для измерения и анализа жевания. Затем обсуждается функциональная роль жевания во время еды и размышлений, подчеркивая его основной вклад в пищеварение и контроль потребления. В последней части этой главы обсуждаются вариации ритма и продолжительности жевательной деятельности в зависимости от характеристик корма и рациона, с одной стороны, и характеристик животных и групп, с другой.
 Авторы):
Белс, В.
Бауссар, С.
Авторы):
Белс, В.
Бауссар, С.
 Авторы):
Поповичс, Т.Е.
Херринг, С.В.
Авторы):
Поповичс, Т.Е.
Херринг, С.В.
 М.
М.
 М.
М.
 Авторы):
Гиденн, Т.
Лебас, Ф.
Авторы):
Гиденн, Т.
Лебас, Ф.
 Д.
Вильяльба, Дж. Дж.
Д.
Вильяльба, Дж. Дж.
 Авторы):
Питтрофф, В.
Сока, П.
Авторы):
Питтрофф, В.
Сока, П.