
Глицин-Био Фармаплант таблетки подъязычн. 100мг 50 шт.
Краткое описание
Препарат, улучшающий метаболизм головного мозга
Состав
1 таблетка содержит:
Активного вещества:
Глицина 0,05 г и 0,1 г.
Вспомогательные вещества:
Повидон (коллидон 25), микрокристаллическая целлюлоза, стеарат магния.
Фармакологическое действие
Средство, влияющее на мозговой метаболизм, заменимая аминокислота. Является центральным нейромедиатором тормозного типа действия. Улучшает метаболические процессы в тканях мозга, оказывает седативное и антидепрессивное действие.
- обладает глицин- и GABA-ергическим, 1-адреноблокирующим, антиоксидантным и антитоксическим действием;
- регулирует деятельность NMDA-рецепторов и за счет этого уменьшает психоэмоциональное напряжение, агрессивность и конфликтность;
- улучшает социальную адаптацию и настроение, облегчает засыпание и нормализует сон;
- повышает умственную работоспособность;
- уменьшает выраженность вегетативно-сосудистых нарушений (в т.
 ч. и в климактерическом периоде) и общемозговых расстройств при ишемическом инсульте и черепно-мозговой травме, токсическое действие этанола на ЦНС.
ч. и в климактерическом периоде) и общемозговых расстройств при ишемическом инсульте и черепно-мозговой травме, токсическое действие этанола на ЦНС. - эффективен в качестве вспомогательного средства при эпилептических приступах.
Показания
В составе комплексной терапии: стрессовые состояния, психоэмоциональное напряжение, снижение умственной работоспособности, нарушения поведения детей и подростков, хронический алкоголизм, прерывание запоя, острый период абстинентного синдрома, функциональные и органические поражения нервной системы, сопровождающиеся повышенной возбудимостью, эмоциональной нестабильностью, снижением умственной работоспособности и нарушением сна (в т.ч. неврозы, неврозоподобные состояния, энцефалопатия различного генеза, остаточные явления после травм головного и спинного мозга), ишемический инсульт.
Способ применения и дозировка
Применяют сублингвально или трансбуккально.
Разовая доза для взрослых — 100-200 мг, для детей (в зависимости от возраста) — 50-100 мг. Кратность приема — 2-3 раза/сут. Длительность применения и частота проведения курсов зависит от показаний и течения заболевания.
Побочные действия
Возможно: аллергические реакции.
Противопоказания
Повышенная чувствительность к глицину.
Особые указания
У пациентов со склонностью к артериальной гипотензии дозы следует уменьшить и контролировать АД, при его уменьшении применение глицина следует отменить.
Взаимодействие с другими препаратами
Уменьшает токсическое действие противосудорожных и антипсихотических средств, антидепрессантов.
При сочетании со снотворными средствами, транквилизаторами и нейролептиками суммируется эффект торможения ЦНС.
Глицин до еды или после
Стрессам подвержены все «от мала до велика». Поэтому препарат Глицин, в состав которого входит аминокислота, важная для умственной деятельности, подходит всем членам семьи. Поскольку препарат популярен, то часто возникает вопрос: принимать Глицин до или после еды? Ведь многое зависит не только от дозировки, но и времени приема средства.
Общее представление о препарате
Приобретая упаковку препарата, вы обнаружите в ней 50 таблеток белого цвета. Каждая таблетка содержит 100мг. глицина, основное составляющее вещество которого – аминоуксусная кислота. Лекарство имеет сладкий вкус, поэтому понравится даже малышу.
Его функция – привести в норму обменные процессы в клетках и тканях мозга. Аминокислота вырабатывается самим организмом, но когда ткани нервной системы поражены, она должна поступать извне.
В небольших дозах никакого вреда организму средство не нанесет, ведь в его составе – природный компонент, поэтому другое его название – пищевая добавка. Но при лечении определенных расстройств нужно знать дозировку и правила приема.
К сведению: Научных доказательств влияния Глицина на работоспособность нет. Это понятно, поскольку аминокислота синтезируется организмом, и отследить ее влияние сложно. Но когда происходят сбои в работе НС, ее влияние неоспоримо.
Инструкция применения и дозы при нервных расстройствах
Как правильно пить Глицин, сказано в инструкции. Вернее, его не пьют, а держат под языком, где он постепенно рассасывается. Людям любого возраста (детям только от 7 лет) назначают одинаковую дозу при следующих симптомах:
- плохой памяти и невнимательности,
- перегрузках эмоционального характера,
- сниженной умственной деятельности,
- замедленном умственном развитие,
- девиантном поведение.

Доза назначается врачом и составляет 1 таблетку 2-3 раза в день. Принимать лекарство следует от 2 недель до месяца. В остальных случаях норма приема будет другая и связана она с конкретным заболеванием. Употребление препарата не связано с приемом пищи, но некоторые врачи советуют его принимать до или во время еды. Во всяком случае, серьезный акцент на этом делать не следует.
Глицин в лечении детей
Средство относят к мягким ноотропным препаратам, который не повредит детям. Когда принимать Глицин ребенку, решают родители. С 2-3 лет норма приема – 0,5 таблетки. От 7 лет дают уже по таблетке БАДа за один прием. Применение трижды в день наиболее оптимально: утром активизируется работа мозга и повышается успеваемость в школе, днем хорошо снимается накопившаяся усталость, а вечером нормализуется сон.
К сведению: Грудной ребенок получает Глицин с молоком матери, которая принимает препарат. Так он лучше усваивается и его легче выпить малышу. Можно в 1⁄4 часть измельченной таблетки обмакнуть соску.
Можно в 1⁄4 часть измельченной таблетки обмакнуть соску.
Обычно детском возрасте лечение назначается на месяц. Как правильно пить Глинин детям: до еды или после еды? Лучшее время — прием до еды или независимо от питания. Отзывы родителей подтверждают эффективность использования Глицина в детском возрасте. Обратите внимание: таблетку нельзя разводить водой, соком или компотом. Так снижается всасывание в кровоток и теряется активность.
Глицин в лечении взрослых
Об общих нормах приема было сказано выше. При поражениях головного мозга используют ту же дозировку. Не меняется она и при алкогольной или наркотической зависимости. Пить лекарство рекомендовано месяц. При продолжительной бессоннице таблетку кладут под язык за полчаса до отхода ко сну.
Доза значительно увеличивается при ишемическом инсульте: через 3-6 часов после инсульта – 10 таблеток под язык, затем по 10 штук каждые 5 дней, после чего назначается обычный прием – 3 раза в день по таблетке.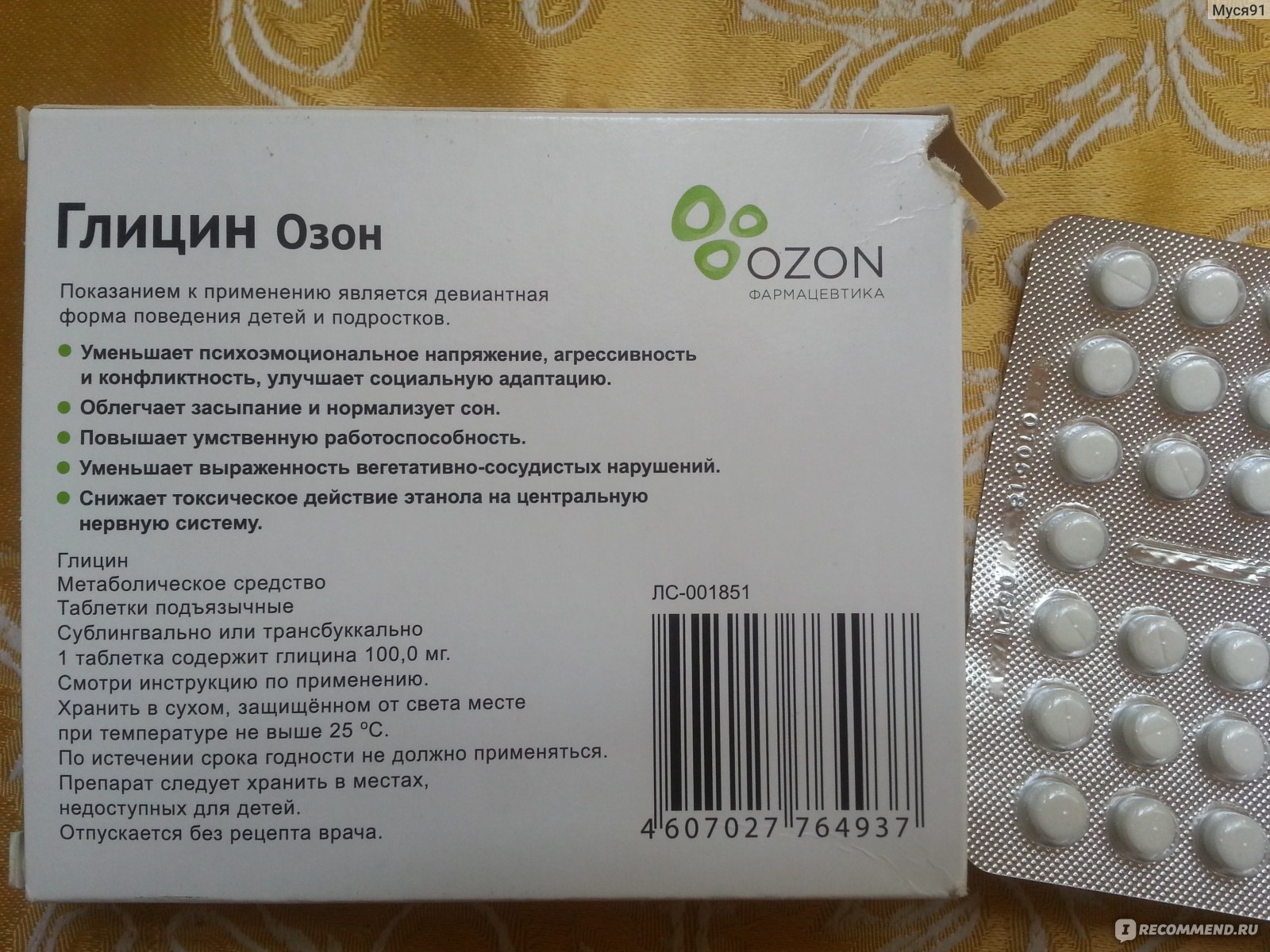 Во время алкогольного опьянения, чтобы вернуть трезвость мыслей, Глицин принимают по 1 таблетке с промежутком в 15-20 мин.
Во время алкогольного опьянения, чтобы вернуть трезвость мыслей, Глицин принимают по 1 таблетке с промежутком в 15-20 мин.
Не придавайте серьезного значения, пить Глицин до еды или после, лучше принимайте его между приемами пищи. Главное, использовать верную дозировку и ждать, что средство поможет пережить еще один трудный день.
Источники:
Видаль: https://www.vidal.ru/drugs/glycine__4429
ГРЛС: https://grls.rosminzdrav.ru/Grls_View_v2.aspx?routingGuid=80b010cc-df9b-4601-90e4-d8cd11a43524&t=
Нашли ошибку? Выделите ее и нажмите Ctrl + Enter
Glycine Глицин 1000 мг, 100 капс NOW foods
Glycine Глицин 1000 мг, 100 капс NOW foods
Одна капсула содержит:
Глицин (аминоуксусная кислота, гликокол) 1000 мг
- Глицин (аминоуксусная кислота, гликокол) влияет на метаболические процессы.
- Глицин является регулятором обмена веществ, нормализует и активирует процессы защитного торможения в центральной нервной системе, уменьшает психоэмоциональное напряжение, повышает умственную работоспособность.

- Глицин метаболизируется до воды и углекислого газа, накопление его в тканях не происходит.
- Глицин является предшественником креатина, который используется для построения ДНК и РНК. Также поддерживает запасы гликогена, в результате чего в организме образуется больше глюкозы для производства энергии.
Функциональное действие:
Глицин обладает глицин- и ГАМК-эргическим, альфа1-адреноблокирующим, антиоксидантным, антитоксическим действием, регулирует деятельность глутаматных (NMDA) рецепторов, за счёт чего препарат:
— Улучшает настроение.
— Облегчает засыпание и нормализует сон.
— Повышает умственную работоспособность. — Уменьшает вегето-сосудистые расстройства, в т.ч. и в климактерическом периоде.
— Уменьшает выраженность общемозговых расстройств при ишемическом инсульте и черепно-мозговых травмах.
— Уменьшает токсическое действие алкоголя.
Показания к применению:
— Глицин назначают практически здоровым детям, подросткам, взрослым для повышения умственной работоспособности.
— При стрессовых ситуациях — психоэмоциональных напряжениях (в период экзаменов, конфликтных и др. ситуациях).
— Функциональных и органических заболеваниях нервной системы (неврозах, неврозоподобных состояниях и вегето-сосудистой дистонии, последствиях нейроинфекций и черепно-мозговых травм,перинатальных и других энцефалопатий, в том числе алкогольного происхождения), сопровождающихся повышенной возбудимостью, эмоциональной нестабильностью, снижением умственной работоспособности, нарушением сна.
— В качестве нейропротекторного препарата глицин назначают при ишемическом инсульте.
Способ применения: по 1 капсуле 1-3 раза в день за 30 мин. до еды.
Практически здоровым детям, подросткам, взрослым для повышения умственной работоспособности, при психоэмоциональных напряжениях, снижении памяти, внимания, задержке умственного развития, при девиантных формах поведения по 1 капсуле 1-3 раза в день, в течении 2-4 недель. Суточная доза 3 г (3 капсулы).
Суточная доза 3 г (3 капсулы).
При функциональных и органических поражениях нервной системы, сопровождающихся повышенной возбудимостью, эмоциональной лабильностью и нарушением сна, детям до 3 лет по 1 капсуле 1-2 раза в день в течении 1-2 недель, в дальнейшем по 1 капсуле 1 раз в день в течении 7-10 дней. Детям старше 3 лет и взрослым по 1 капсуле 2-3 раза в день в течении 1-2 недель. При необходимости курс лечения повторяют.
При нарушении сна глицин назначают за 20 минут до сна или непосредственно перед сном по 1 капсуле (в зависимости от возраста).
При ишемическом мозговом инсульте: в течении первых 3-6 часов по 1капсуле с одной чайной ложкой воды, далее в течении 5 суток по 3 капсулы в сутки, затем в течении последующих 20 дней по 1 капсуле 3 раза в сутки. В наркологии глицин применяют в качестве средства, повышающего умственную работоспособность и уменьшающего психоэмоциональное напряжение в период ремиссии при явлениях энцефалопатии, органических поражениях центральной и периферической нервной системы по 1 капсуле 2-3 раза в сутки в течении 2-4 недель. При необходимости курсы повторяют 4-6 раз в год (каждые 2-3 месяца).
При необходимости курсы повторяют 4-6 раз в год (каждые 2-3 месяца).
Побочное действие: возможны аллергические реакции.
Противопоказания: индивидуальная непереносимость.
Условия хранения: в сухом, прохладном, недоступном для детей месте, t° не выше +25°С.
Правила подготовки к лабораторным исследованиям
Исследование крови
Показатели крови могут существенно меняться в течение дня, поэтому все анализы следует сдавать в утренние часы. Все анализы крови следует сдавать до проведения рентгенографии, УЗИ, ФГДС и физиотерапевтических процедур. В течение часа до сдачи крови необходимо воздержаться от курения. За сутки до сдачи крови желательно избегать физических нагрузок, приема алкоголя и существенных изменений в питании и режиме дня. На результаты исследований может влиять прием лекарственных препаратов. Если Вы принимаете лекарства, обязательно предупредите об этом лечащего врача и процедурную медсестру. При необходимости исследования натощак, между последним приемом пищи и взятием крови должно пройти не менее 12 часов. Можно пить воду и принимать лекарства. Если ранее при взятии крови Вы испытывали головокружение, предупредите заранее процедурную сестру — кровь у Вас возьмут в положении лежа.
При необходимости исследования натощак, между последним приемом пищи и взятием крови должно пройти не менее 12 часов. Можно пить воду и принимать лекарства. Если ранее при взятии крови Вы испытывали головокружение, предупредите заранее процедурную сестру — кровь у Вас возьмут в положении лежа.
ВАЖНО! После взятия крови не рекомендуется в течение 1 часа нагружать руку, из вены которой проводили взятие крови (например, нести сумку).
При контроле лабораторных показателей в динамике рекомендуется проводить повторные исследования в одинаковых условиях: в одной лаборатории, сдавать кровь в одинаковое время суток и пр. Взятие венозной крови производится с помощью вакуумных закрытых систем. В вену будет введена игла и не исключено, что во время прокола Вы можете испытать кратковременное незначительное болевое ощущение. После взятия крови иглу удалят. Крайне редко, после взятия крови на месте прокола может возникнуть гематома. Особой опасности данный вид гематомы не представляет. Чтобы она не появилась, надо четко следовать советам медицинского персонала. С целью профилактики образования гематомы (синяка) на место прокола накладывается тугая повязка с использованием стерильного материала. Удаление повязки возможно через 15 минут. Любой другой метод менее эффективен для профилактики образования гематомы.
Чтобы она не появилась, надо четко следовать советам медицинского персонала. С целью профилактики образования гематомы (синяка) на место прокола накладывается тугая повязка с использованием стерильного материала. Удаление повязки возможно через 15 минут. Любой другой метод менее эффективен для профилактики образования гематомы.
Клинический анализ крови
Кровь для исследования чаще берут из вены, реже из пальца, у новорожденного — из пятки. Если назначен только общий анализ крови, то последний прием пищи — за час до сдачи крови.
Биохимический анализ крови, исследование гормонов крови, коагулограмма
Исследуют венозную кровь натощак. Между последним приемом пищи и взятием крови должно пройти не менее 12 часов. Можно пить воду и принимать лекарства.
ВАЖНО! При исследовании липидограммы (холестерин и фракции) — между последним приемом пищи (легкий ужин) и взятием крови должно пройти не менее 14 часов. Можно пить воду и принимать лекарства.
Можно пить воду и принимать лекарства.
При проведении тестов толерантности к глюкозе в течение 3 дней следует соблюдать смешанную диету, содержащую свыше 250 гр углеводов в день (обычный рацион ).
ВАЖНО! Необходимо согласовать режим приема медикаментов, способных влиять на обмен глюкозы с врачом. Длительность проведения теста в зависимости от назначений врача может быть от 1 часа до 4 часов. Это время пациент должен находиться в клинике.
ВАЖНО! При исследовании гормонального статуса женщинам репродуктивного возраста следует обратить особое внимание на необходимость сдачи крови в соответствующий день менструального цикла.
ВАЖНО! При исследовании коагулограммы обязательно сообщить процедурной медсестре принимаемые лекарственные препараты (особенно важно варфарин, гепарин, фраксипарин, клексан, плавикс, аспирин).
Остальные исследования крови не требуют строгих ограничений.
Исследование мокроты
Мокроту собирают утром, до приема пищи, в пластиковые одноразовые стерильные контейнеры. Контейнер можно получить на приеме у врача или приобрести в аптеке. Перед откашливанием мокроты почистить зубы. Рот прополоскать водой. Вымыть руки. Сделать несколько глубоких вдохов, после чего энергично покашлять и выплюнуть мокроту в контейнер. Закрыть крышку.
Контейнер можно получить на приеме у врача или приобрести в аптеке. Перед откашливанием мокроты почистить зубы. Рот прополоскать водой. Вымыть руки. Сделать несколько глубоких вдохов, после чего энергично покашлять и выплюнуть мокроту в контейнер. Закрыть крышку.
Общий анализ мочи и анализ мочи по Нечипоренко
Перед сбором мочи обязательно проводят тщательный туалет наружных половых органов. Для исследования собирают утреннюю (сразу после сна) среднюю порцию мочи в емкость.
ВНИМАНИЕ! Первую и последнюю порции мочи выпускают в унитаз.
Для взятия используют шприц — пробирки желтого цвета.
См. схему.
При одновременном назначении общего анализа мочи и мочи по Нечипоренко, необходимо не менее 2 пробирок по 12 мл. При использовании пластикового контейнера мочу собрать обычным способом. При одновременном назначении общего анализа мочи и мочи по Нечипоренко, необходимо не менее 25 мл мочи.
Внимание! Моча доставляется в лабораторию в день взятия. До следующего дня ее хранить нельзя.
Моча, собранная для общего анализа, может храниться не более 1,5 — 2 ч (обязательно в прохладном месте, например холодильнике при температуре 2-8°С). Нельзя собирать мочу во время менструации и в течение 2-3 дней по окончании, в течение 5-7 дней после цистоскопии.
Бактериологическое исследование мочи
Используется пробирка зеленого цвета с наполнителем (содержит борную кислоту, которая препятствует размножению бактерий). Техника взятия мочи — см. общий анализ мочи. Борная кислота стабилизирует рост микроорганизмов в течение 48 часов при хранении пробы мочи при комнатной температуре.
Сбор суточной мочи
Для сбора суточной мочи в аптеке клиники «Скандинавия» можно приобрести специальные наборы, комплектация набора зависит от вида исследования. В каждый набор входит пластиковая емкость 3 литра с мерной шкалой для фиксации суточного объема мочи, пробирки SARSTEDT 12 мл (URINE) желтого цвета и/или и пластиковые одноразовые контейнеры SARSTEDT, в набор для сбора на адреналин, норадреналин, дофамин, ВМК, кальций, фосфор, входит пластиковая пробирка со стабилизатором. Перед началом сбора мочи содержимое пробирки вылить в 3-х литровую емкость. Мочу собирают в течение 24 часов в один большой контейнер (банку). Последняя порция мочи берется точно в то же время, когда был начат сбор. Измеряют объем мочи (суточный диурез) в миллилитрах (мл). Перемешивают. В лабораторию для исследования доставляют примерно 50 мл. В направлении указывают объем выделенной мочи за сутки (суточный диурез) в миллилитрах (мл).
Перед началом сбора мочи содержимое пробирки вылить в 3-х литровую емкость. Мочу собирают в течение 24 часов в один большой контейнер (банку). Последняя порция мочи берется точно в то же время, когда был начат сбор. Измеряют объем мочи (суточный диурез) в миллилитрах (мл). Перемешивают. В лабораторию для исследования доставляют примерно 50 мл. В направлении указывают объем выделенной мочи за сутки (суточный диурез) в миллилитрах (мл).
Результаты анализов в вашем мобильном приложении
Подробнее >>>
Суточная экскреция (потеря) белка , 17-КС, 17-ОК
Для сбора суточной мочи в аптеке клиники «Скандинавия» можно приобрести специальный набор. В течение суток соблюдают обычный питьевой режим. Утром в 6-8 часов освобождают мочевой пузырь (эту порцию выливают). Далее собирают мочу в течение 24 часов в один большой контейнер (банку).
Далее собирают мочу в течение 24 часов в один большой контейнер (банку).
ВНИМАНИЕ! Моча не нуждается в хранении в холодном месте.
Последняя порция берется точно в то же время, когда был начат сбор. Измеряют объем мочи (суточный диурез) в миллилитрах (мл). Перемешивают. В лабораторию для исследования доставляют примерно 50 мл. В направлении указывают объем выделенной мочи за сутки (суточный диурез) в миллилитрах (мл).
Правила сбора мочи за сутки на: адреналин, норадреналин, кальций, фосфор, ванилилминдальная, гомованилиновая и 5-гидроксииндолуксусная кислоты, метанефрин и норметанефрин
Для сбора суточной мочи на эти исследования в аптеке клиники «Скандинавия» можно приобрести специальный набор с консервантом. В течение суток соблюдают обычный питьевой режим. Утром в 6-8 часов освобождают мочевой пузырь (эту порцию выливают). Следующую порцию мочи собирают в большую емкость и туда же выливают содержимое флакона со стабилизатором. Далее собирают мочу в течение 24 часов в один большой контейнер (банку). Последняя порция мочи берется точно в то же время, когда был начат сбор. Измеряют объем мочи (суточный диурез) в миллилитрах (мл). Перемешивают. В лабораторию для исследования доставляют примерно 50 мл. В направлении указывают объем выделенной мочи за сутки (суточный диурез) в миллилитрах (мл).
Далее собирают мочу в течение 24 часов в один большой контейнер (банку). Последняя порция мочи берется точно в то же время, когда был начат сбор. Измеряют объем мочи (суточный диурез) в миллилитрах (мл). Перемешивают. В лабораторию для исследования доставляют примерно 50 мл. В направлении указывают объем выделенной мочи за сутки (суточный диурез) в миллилитрах (мл).
Анализ кала: копрограмма, яйца глистов, простейшие, реакция Грегерсена («скрытая кровь»)
Кал доставляют в пластиковом контейнере (объем материала — с грецкий орех). При одновременном исследовании кала на яйца глистов и копрограмму необходимо 2 контейнера. При назначении реакции Грегерсена «скрытая кровь» необходимо строго придерживаться диеты в течение 3 суток и исключить из пищи рыбу, мясо, зеленые овощи, помидоры, а также препараты железа. Кал должен быть доставлен для исследования не позднее, чем через 12 часов после дефекации при условии хранения холодильнике (+2 — +8°С). Цисты лямблий стабильны, поэтому для обнаружения цист срок доставки — до 24 часов.
Цисты лямблий стабильны, поэтому для обнаружения цист срок доставки — до 24 часов.
Анализ кала на дисбактериоз
Обязательное условие — утренняя порция кала. При невозможности собрать анализ в это время посоветуйтесь с врачом. Кал доставляют в специальной пластиковой емкости для кала SARSTEDT (объем материала — с грецкий орех). Контейнер доставляют в лабораторию не позднее 2 часа после сбора пробы или при условии хранения в холодильнике 4 часа (при температуре 4-8 °С).
Уточнить время приема анализов можно по телефону нашего колл-центра, а результаты всегда можно увидеть в личном кабинете или мобильном приложении.
Amazon.com: NOW Supplements, Glycine Pure Powder, способствует спокойному сну *, поддержка нейротрансмиттеров *, 1 фунт: Health & Household
NOW Supplements Glycine Pure Powder обеспечит вам безопасный и спокойный сон. Также он оказывает сильное успокаивающее действие. Это вместе с мощным набором пищевых добавок, которые я принимаю, изменило качество моей жизни.
Это вместе с мощным набором пищевых добавок, которые я принимаю, изменило качество моей жизни. Теперь я сплю дольше и глубже. Бессонница значительно улучшилась. Беспокойство не только уменьшилось, но и ушло. Я больше не страдаю депрессией, ОКР значительно улучшилось, фокус улучшился, мозг рассеялся.Я теряю вес, похудела на 2,5 дюйма ниже талии, и все эти эффекты приписываю глицину в дополнение к пачке добавок, которую я сейчас принимаю. Предиабет и инсулинорезистентность ушли. АД в норме. Здоровые уровни глюкозы / AC1 / HDL / LDL в моем анализе крови. Никаких изменений в питании и физических упражнениях, но я выгляжу и чувствую себя на 20 лет моложе. У меня есть энергия! Мой доктор удивлен. 6 недель — все, что мне потребовалось.
Этот порошок глицина чистый, не имеет нежелательного вкуса и с ним очень легко разговаривать. Я добавляю его в кофе с обезжиренными сливками и сливками из лесных орехов, и считаю, что это не меняет вкуса.Мне нравится, что это чистый глицин, не смешанный с добавками, которые я нахожу в форме капсул. А так как это порошок, вы можете легко взять точное количество, которое хотите, так что вы сами контролируете дозировку.
А так как это порошок, вы можете легко взять точное количество, которое хотите, так что вы сами контролируете дозировку.
Мне пришлось столкнуться с длительным трудным периодом моей жизни. Добавьте к этому, что я страдаю хронической бессонницей, и из-за тяжелого посттравматического стрессового расстройства я страдаю от постоянного нахождения в состоянии повышенной бдительности, с напряженными мышцами и ловлю себя на сжатых руках и напряженных плечах / шее. Пытаться лечь и заснуть так невозможно.Пытаться просто нормально функционировать в течение дня было невозможно. Пытаться работать на двух-трехчасовом сне невозможно. К тому же все это вызывает высокий уровень кортизола и адреналина; Я смотрел, как расширяется моя талия. Мой холестерин взлетел до небес, кровяное давление резко возросло, и мне поставили диагноз «преддиабет».
Мой терапевт больше года приставал ко мне, чтобы я попробовал рецепт, и когда я наконец отступил от отчаяния, лекарства, отпускаемые по рецепту, ухудшили мои симптомы, а также у меня появилось несколько новых, включая пугающую дрожь рук. В первую неделю выбросил эти лекарства в мусорное ведро. Я начал проводить исследования, попробовал несколько комбинаций натуральных добавок, которые не помогли, и, наконец, нашел комбинацию, которая работает. Глицин — важная и неотъемлемая часть естественного, клинически подтвержденного сочетания, которое изменило мою жизнь.
В первую неделю выбросил эти лекарства в мусорное ведро. Я начал проводить исследования, попробовал несколько комбинаций натуральных добавок, которые не помогли, и, наконец, нашел комбинацию, которая работает. Глицин — важная и неотъемлемая часть естественного, клинически подтвержденного сочетания, которое изменило мою жизнь.
Я обычно добавляю этот порошок глицина в свой утренний кофе, а затем снова добавляю немного в напиток ранним вечером. Я сразу чувствую спокойствие и ощущаю настоящее расслабление.
Характеристика интерактивных эффектов глицина и D-циклосерина у мужчин: дополнительные доказательства улучшенной функции рецептора NMDA, связанной с алкогольной зависимостью человека
Allgaier C, Scheibler P, Muller D, Feuerstein TJ, Illes P (1999).Характеристика рецепторов NMDA и экспрессия субъединиц в культивируемых мезэнцефальных нейронах крыс. Br J Pharmacol 126 : 121–130.
CAS Статья PubMed PubMed Central Google ученый
Барнетт Дж. Х., Салмонд СН, Джонс ПБ, Саакян Б. Дж. (2006). Когнитивный резерв в нейропсихиатрии. Psychol Med 36 : 1053–1064.
Х., Салмонд СН, Джонс ПБ, Саакян Б. Дж. (2006). Когнитивный резерв в нейропсихиатрии. Psychol Med 36 : 1053–1064.
CAS Статья PubMed Google ученый
Barr LC, D’Souza DC, Price LH, Krystal JH (1994).Оценка дозозависимых эффектов D-циклосерина у пациентов с обсессивно-компульсивным расстройством без лечения. Soc Neurosci Abstr 20 : 1564 (номер аннотации 641.9).
Google ученый
Bienkowski P, Krzascik P, Koros E, Kostowski W, Scinska A, Danysz W (2001). Влияние нового неконкурентоспособного антагониста рецептора NMDA, MRZ 2/579, на самовведение этанола и судороги его отмены у крыс. Eur J Pharmacol 413 : 81–89.
CAS Статья PubMed Google ученый
Бьенковски П., Стефански Р., Костовски В. (1997). Дискриминационные стимулирующие эффекты этанола: отсутствие антагонизма с N -метил-D-аспартатом и D-циклосерином. Спирт 14 : 345–350.
CAS Статья PubMed Google ученый
Бисага А., Эванс С.М. (2004).Острые эффекты мемантина в сочетании с алкоголем у умеренно пьющих. Психофармакология 172 : 16–24.
CAS Статья PubMed Google ученый
Брандт Дж. (1991). Тест на вербальное обучение Хопкинса: разработка нового теста на память с шестью эквивалентными формами. Клинический нейропсихолог 5 : 125–142.
Артикул Google ученый
Бремнер Дж. Д., Кристал Дж. Х., Патнэм Ф. У., Саутвик С. М., Мармар К., Чарни Д. С. и др. (1998).Измерение диссоциативных состояний с помощью шкалы диссоциативных состояний, управляемой клиницистом (CADSS). J Стресс травмы 11 : 125–136.
CAS Статья Google ученый
Бруннер Э, Домхоф С., Лангер Ф (2002). Непараметрический анализ продольных данных в факторных экспериментах . Wiley: Хобокен, штат Нью-Джерси.
Google ученый
D’Souza DC, Gil R, Cassello K, Morrissey K, Abi-Saab D, White J et al (2000).Влияние глицина и D-циклосерина внутривенно на аминокислоты в плазме и спинномозговой жидкости у здоровых людей. Биологическая психиатрия 47 : 450–462.
Артикул PubMed Google ученый
Даниш В., Парсонс К.Г. (1988). Рецепторы глицина и N -метил-D-аспартата: физиологическое значение и возможные терапевтические применения. Pharmacol Rev 50 : 597–664.
Google ученый
Дарштайн М.Б., Ландвермейер ГБ, Фейерштейн Т.Дж. (2000).Изменения экспрессии гена субъединицы рецептора NMDA в головном мозге крысы после отказа от принудительного длительного приема этанола. Naunyn Schmiedebergs Arch Pharmacol 361 : 206–213.
CAS Статья PubMed Google ученый
Давид В., Ионеску М., Думитреску В. (2001). Определение циклосерина в плазме человека с помощью высокоэффективной жидкостной хроматографии с детектированием флуоресценции с использованием дериватизации с использованием p -бензохинона. J Chromatogr B 761 : 27–33.
CAS Статья Google ученый
Дикерсон Д., Питтман Б., Ралевски Е., Перрино А., Лимончелли Д., Эджкомб Дж. и др. (2010). Этанолоподобные эффекты тиопентала и кетамина у здоровых людей. J Psychopharmacol 24 : 203–211.
CAS Статья PubMed Google ученый
Эмметт М.Р., Мик С.Дж., Клер Дж.А., Рао Т.С., Айенгар С., Вуд П.Л. (1991).Действия D-циклосерина на N -метил-D-аспартат-ассоциированный сайт рецептора глицина in vivo . Нейрофармакология 30 : 1167–1171.
CAS Статья PubMed Google ученый
Эванс С.М., Левин Ф.Р., Брукс Д.Д., Гарави Ф. (2007). Пилотное двойное слепое испытание лечения алкогольной зависимости мемантином. Alcohol Clin Exp Res 31 : 775–782.
CAS Статья PubMed Google ученый
Фейн Г., МакГилливрей С. (2007).Когнитивные способности у пожилых алкоголиков, длительное время воздерживающихся от употребления алкоголя. Alcohol Clin Exp Res 31 : 1788–1799.
Артикул PubMed PubMed Central Google ученый
Гонсалес Р.А., Браун Л.М. (1995). Региональные различия в мозговом реверсировании глицином индуцированного этанолом ингибирования N -метил-D-аспартат-стимулированного высвобождения нейротрансмиттера. Life Sci 56 : 571–577.
CAS Статья PubMed Google ученый
Гордон М. (1983). Гордон Диагностические Системы . Системы Гордона: ДеВитт, штат Нью-Йорк.
Google ученый
Грант К.А., Ловингер Д.М. (1995). Клеточная и поведенческая нейробиология алкоголя: нейрональные процессы, опосредованные рецепторами. Clin Neurosci 3 : 155–164.
CAS PubMed Google ученый
Харихаран М., Нага С., Ванноорд Т. (1993). Системный подход к развитию аминокислотного анализа плазмы с помощью высокоэффективной жидкостной хроматографии с ультрафиолетовым детектированием с предколоночной дериватизацией с использованием фенилизотиоцианата. J Chromatogr 621 : 15–22.
CAS Статья PubMed Google ученый
Хоффман П.Л. (2003 г.). Рецепторы NMDA при алкоголизме. Int Rev Neurobiol 56 : 35–82.
CAS Статья PubMed Google ученый
Хоффман П.Л., Грант К.А., Снелл Л.Д., Рейнлиб Л., Иорио К., Табакофф Б. (1992). Рецепторы NMDA: роль в припадках отмены этанола. Ann N Y Acad Sci 654 : 52–60.
CAS Статья PubMed Google ученый
Hood WF, Compton RP, Monahan JB (1989). -Циклосерин: лиганд N -метил-D-аспартат-связанный глициновый рецептор имеет характеристики частичного агониста. Neurosci Lett 98 : 91–95.
CAS Статья PubMed Google ученый
Коллер Г., Зилл П., Фер С., Погарелл О., Бонди Б., Сойка М. и др. (2009).Отсутствие ассоциации алкогольной зависимости с полиморфизмами переносчиков глицина SLC6A5 и SLC6A9. Addict Biol 14 : 506–508.
CAS Статья PubMed Google ученый
Kostowski W, Bienkowski P (1999). Дискриминационные стимульные эффекты этанола: нейрофармакологическая характеристика. Спирт 17 : 63–80.
CAS Статья PubMed Google ученый
Kotlinska J, Liljequist S (1997).Антагонист рецептора NMDA / глицина, L-701,324, производит дискриминационные стимулы, аналогичные стимулам этанола. Eur J Pharmacol 332 : 1–8.
CAS Статья PubMed Google ученый
Крюгер Дж. М., О’Коннор А. Дж., Келсо С. Р. (1997). Агонисты глицинового сайта проявляют субъединично-специфические эффекты на рецепторы NMDA, экспрессируемые в ооцитах Xenopus . Soc Neurosci Abstr 23 : 945.
Google ученый
Крупицкий Е.М., Незнанова О., Масалов Д., Бураков А.М., Диденко Т., Романова Т. и др. (2007а). Влияние мемантина на вызванную сигналом тягу к алкоголю у выздоравливающих пациентов с алкогольной зависимостью. Am J Psychiatry 164 : 519–523.
Артикул PubMed Google ученый
Крупицкий Е.М., Руденко А.А., Бураков А.М., Славина Т.Ю., Гриненко А.А., Питтман Б и др. (2007б).Антиглутаматергические стратегии детоксикации этанолом: сравнение с плацебо и диазепамом. Alcohol Clin Exp Res 31 : 604–611.
CAS PubMed Google ученый
Кристал Дж., Петракис И., Красницки С., Тревизан Л., Бутрос Н., Д’Суза, округ Колумбия (1998a). Измененные ответы на агонисты нечувствительного к стрихнину сайта коагониста глицина NMDA (SIGLY) у недавно прошедших детоксикацию алкоголиков. Alcohol Clin Exp Res 22 : 94A.
Google ученый
Кристал Дж. Х., Петракис И. Л., Лимончелли Д., Уэбб Е., Георгуева Р., Д’Суза, округ Колумбия, и др. (2003a). Измененный ответ антагониста рецептора глутамата NMDA у выздоравливающих пациентов, зависимых от этанола. Нейропсихофармакология 28 : 2020–2028.
CAS Статья PubMed Google ученый
Кристал Дж. Х., Петракис Иллинойс, Мейсон Дж., Д’Суза, округ Колумбия (2003b).Рецепторы глутамата NMDA и алкоголизм: награда, зависимость, лечение и уязвимость. Pharmacol Therap 99 : 79–94.
CAS Статья Google ученый
Кристал Дж. Х., Петракис Иллинойс, Тревизан Л., Д’Суза, округ Колумбия (2003c). Антагонизм рецепторов NMDA и сигнал интоксикации этанолом: от риска алкоголизма к фармакотерапии. Ann N Y Acad Sci 1003 : 176–184.
CAS Статья PubMed Google ученый
Кристал Дж. Х., Петракис И. Л., Уэбб Э., Куни Н. Л., Карпер Л. П., Наманворт С. и др. (1998b).Дозозависимые этанолоподобные эффекты антагониста NMDA, кетамина, у недавно прошедших детоксикацию алкоголиков. Arch Gen Psychiatry 55 : 354–360.
CAS PubMed Google ученый
Кристал Дж. Х., Петракис И. Л., Уэбб Э., Куни Н. Л., Карпер Л. П., Наманворт С. и др. (1998c). Дозозависимые этанолоподобные эффекты антагониста NMDA, кетамина, у недавно прошедших детоксикацию алкоголиков. Arch Gen Psychiatry 55 : 354–360.
CAS PubMed Google ученый
Кристал Дж. Х., Уэбб Э., Куни Н., Кранцлер Х. Р., Чарни Д. С. (1994). Специфика этанолоподобных эффектов, вызываемых серотонинергическими и норадренергическими механизмами. Архив общей психиатрии 51 : 898–911.
CAS Статья Google ученый
Кумари М., Тику МК (2000). Регулирование рецепторов NMDA этанолом. Progr Drug Res 54 : 151–189.
CAS Статья Google ученый
Lido HH, Стомберг Р., Фагерберг А., Эриксон М., Содерпалм Б. (2009). Ингибитор обратного захвата глицина org 25935 взаимодействует с базальным и индуцированным этанолом высвобождением дофамина в прилежащем ядре крысы. Alcohol Clin Exp Res 33 : 1151–1157.
Артикул PubMed Google ученый
Мартин С.С., Эрливайн М., Мусти Р.Э., Перрин М.В., Свифт Р.М. (1993).Разработка и проверка шкалы двухфазных эффектов алкоголя. Alcohol Clin Exp Res 17 : 140–146.
CAS Статья PubMed Google ученый
Моландер А, Лидо ХХ, Лоф Э, Эриксон М, Содерпалм В (2007). Ингибитор обратного захвата глицина Org 25935 снижает потребление этанола и его предпочтение у самцов крыс линии Вистар. Алкоголь Спирт 42 : 11–18.
CAS Статья PubMed Google ученый
Морган С.Дж., Перри Э.Б., Чо Х.С., Кристал Дж. Х., Д’Суза, округ Колумбия (2006).Повышенная уязвимость к амнестическим эффектам кетамина у мужчин. Психофармакология (Берл) 187 : 405–414.
CAS Статья Google ученый
Петракис И.Л., Лимончелли Д., Георгиева Р., Джатлоу П., Бутрос Н.Н., Тревизан Л. и др. (2004). Измененный ответ антагониста рецептора глутамата NMDA у лиц с семейной уязвимостью к алкоголизму. Am J Psychiatry 161 : 1776–1782.
Артикул PubMed Google ученый
Rammes G, Mahal B, Putzke J, Parsons C, Spielmanns P, Pestel E et al (2001). Соединение акампросат против тяги действует как слабый антагонист NMDA-рецептора, но модулирует экспрессию субъединицы NMDA-рецептора, подобно мемантину и MK-801. Нейрофармакология 40 : 749–760.
CAS Статья PubMed Google ученый
Шутц К.Г., Сойка М (2000).Проблема с декстрометорфаном у пациентов с алкогольной зависимостью и контрольной группы. Arch Gen Psychiatry 57 : 291–292.
CAS Статья PubMed Google ученый
Шейнин А., Шавит С., Бенвенисте М. (2001). Субъединичная специфичность и механизм действия частичного агониста NMDA D-циклосерина. Нейрофармакология 41 : 151–158.
CAS Статья PubMed Google ученый
Спитцер Р.Л., Уильямс Дж. Б., Гиббон М., Первый МБ (1992).Структурированное клиническое интервью для DSM-III-R (SCID). I: История, обоснование и описание. Arch Gen Psychiatry 49 : 624–629.
CAS Статья PubMed PubMed Central Google ученый
Spitzer RL, Williams JBW, Гиббон М., Первый МБ (1990). Структурированное клиническое интервью для DSM-III-R-Nan-Patient Edition (SCID-NP, версия 1.0 с дополнением к DSM-IV) . Американская психиатрическая пресса: Вашингтон, Д.C ..
Google ученый
Стерн Р.А., Сильва С.Г., Чейссон Н., Эванс Д.Л. (1996). Влияние когнитивного резерва на нейропсихологическое функционирование при бессимптомной инфекции вируса иммунодефицита человека-1. Arch Neurol 53 : 148–153.
CAS Статья PubMed Google ученый
Trevisan L, Petrakis IL, Pittman B, Gueorguieva R, D’Souza DC, Perry E et al (2008).Отсутствие значительных интерактивных эффектов высоких доз D-циклосерина и этанола у здоровых людей: предварительные сведения о действиях этанола на глициновом (B) сайте рецепторов глутамата NMDA. Alcohol Clin Exp Res 32 : 36–42.
CAS Статья PubMed Google ученый
фон Кнорринг А.Л., Бохман М., фон Кнорринг Л., Орланд Л. (1985). Активность МАО тромбоцитов как биологический маркер в подгруппах алкоголизма. Acta Psychiatr Scand 72 : 51–58.
CAS Статья PubMed Google ученый
Watson GB, Bolanowski MA, Baganoff MP, Deppeler CL, Lanthorn TH (1990). -Циклосерин действует как частичный агонист на участке модуляции глицина рецептора NMDA, экспрессируемого в ооцитах Xenopus . Brain Res 510 : 158–160.
CAS Статья PubMed Google ученый
Witkin JM, Brave S, French D, Geter-Douglass B (1995).Дискриминантные стимулирующие эффекты R — (+) — 3-амино-1-гидроксипирролид-2-он, [(+) — HA-966], частичного агониста нечувствительного к стрихнину модуляторного сайта N — Рецептор метил-D-аспартата. J Pharmacol Exp Ther 275 : 1267–1273.
CAS PubMed Google ученый
L — ГЛИЦИН — ПИТАНИЕ АТФ
Пищевая добавка
— это аминокислота, которая оказывает успокаивающее действие на мозг и является эффективным средством лечения различных заболеваний, от расстройства желудка до шизофрении.Кроме того, глицин может помочь удалить токсичные вещества, такие как молочная кислота, из организма, а также может увеличить использование и доступность гормона роста. Было доказано, что он способствует высвобождению гормона роста и поддерживает увеличение объема клеток. Когда глицин входит в сбалансированную формулу аминокислот, он помогает замедлить распад мышечной ткани. Это также полезно для заживления поврежденных мышц или простого восстановления мышечной ткани у спортсменов, которые участвуют в экстремальных тренировках.Кроме того, кажется, что глицин увеличивает силу и развитие мышц.
| Информация о пищевой ценности | 1 капсула | 2 капсулы |
| L-глицин | 500 мг * | 1 000 мг * |
* Суточная доза не установлена
Состав: L-глицин, твердая желатиновая капсула, средство против слеживания: фосфат кальция, стеарат магния.
Указания по применению: 2 капсулы 1-2 раза в день между приемами пищи. Пить много воды.
Предупреждения: Не подходит для детей, беременных и кормящих женщин. Хранить в недоступном для детей месте. После открытия хранить в сухом месте при температуре ниже 25 ° C, тщательно закрывать. Производитель не несет ответственности за ущерб, вызванный неправильным использованием или хранением. Нет заменителя разнообразного питания. Не превышайте рекомендуемую дозировку.
В комплекте: 200 капсул (± 5%)
ГЛИЦИН
ГЛИЦИННомер MSDS: G5828 * * * * * Дата вступления в силу: 14.05.03 * * * * * Заменяет: 02.11.01
1.Идентификация продукта
Синонимы: Гликокол, Аминоуксусная кислота
Номер CAS: 56-40-6
Молекулярный вес: 75,07
Химическая формула: Ch3Nh3 COOH
Коды продуктов:
J.T. Бейкер: 0581, 0582, 1479, 4057, 4059
Маллинкродт: 5104, 7728
2. Состав / информация о компонентах
Ингредиент CAS, неопасный в процентах --------------------------------------- ----------- - ------------ --------- Глицин 56-40-6 90-100% Да
3.Идентификация опасностей
Обзор аварийной ситуации
—————————
Как часть правил производственной и личной гигиены и техники безопасности, избегайте всего ненужного воздействия химического вещества и обеспечить быстрое удаление из кожа, глаза и одежда.J.T. Бейкер SAF-T-DATA ™ Рейтинги (указаны здесь для вашего удобства)
————————————————- ————————————————— ———
Рейтинг здоровья: 1 — Незначительный
Рейтинг воспламеняемости: 1 — Незначительный
Рейтинг реактивности: 0 — Нет
Рейтинг контактов: 0 — Нет
Лаборатория Защитное снаряжение: ОЧКИ; LAB COAT
Цветовой код хранения: Оранжевый (Общие) Место хранения)
————————————————- ————————————————— ———Возможное воздействие на здоровье
———————————-Вдыхание:
Высокая концентрация пыли может вызвать кашель.
Проглатывание:
Большие пероральные дозы могут вызвать тошноту.
Скин Обращаться:
Побочных эффектов не ожидается.
Попадание в глаза:
Май вызвать покраснение.
Хроническое воздействие:
Информация не найдена.
Ухудшение ранее существовавших условий:
Информация не найдена.
4. Меры первой помощи
Вдыхание:
Вынести на свежий воздух.Обратитесь за медицинской помощью при любом затруднении дыхания.
Проглатывание:
Дайте несколько стаканов воды выпить, чтобы разбавить. При проглатывании больших количеств обратитесь к врачу. совет.
Контакт с кожей:
Вымойте пораженные участки водой с мылом. При появлении раздражения обратитесь за медицинской помощью.
Попадание в глаза:
Мыть тщательно проточной водой. При появлении раздражения обратитесь за медицинской помощью.
5.Противопожарные мероприятия
Пожар:
Не считается пожароопасным.
Взрыв:
Не считается взрывоопасным.
Средства пожаротушения:
Используйте любые средства, подходящие для тушение окружающего пожара.
Особая информация:
В в случае пожара носить полную защитную одежду и одобрено NIOSH автономный дыхательный аппарат с полнолицевой маской, работающий в требование давления или другой режим положительного давления.
6. Меры при случайной утечке
Неопасный материал. Промыть водой, чтобы канализация.
7. Обращение и хранение
Хранить в плотно закрытой таре, в прохладном, сухом, защищенном от света месте. вентилируемая зона. Защищать от физического повреждения.
8. Контроль воздействия / индивидуальная защита
Пределы воздействия в воздухе:
Не установлены.
Вентиляционная система:
В общем, разбавляющая вентиляция удовлетворительный контроль опасности для здоровья этого вещества.Однако если условия использования создают дискомфорт для рабочего, местная вытяжная система должна быть считается.
Персональные респираторы (одобрено NIOSH):
Для условия использования при очевидном воздействии пыли или тумана и инженерных контроль невозможен, респиратор для твердых частиц (NIOSH тип N95 или лучше фильтры) можно носить. Если частицы масла (например, смазки, смазочно-охлаждающие жидкости, глицерин и т. д.), используйте фильтр NIOSH типа R или P.В экстренных случаях или в случаях, когда уровни воздействия неизвестны, используйте анфас респиратор с подачей воздуха и положительным давлением. ВНИМАНИЕ! Воздухоочистительные респираторы. не защищают рабочих в атмосфере с дефицитом кислорода.
Скин Защита:
Пользоваться защитными перчатками и чистой закрывающей тело одеждой.
Защита глаз:
Защитные очки. Поддерживайте фонтан для промывания глаз и средства для быстрого слива в рабочей зоне.
9.Физико-химические свойства
Внешний вид:
Белые кристаллы.
Запах:
Без запаха.
Растворимость:
25 г / 100 мл воды при 25 ° C (77 ° F).
Плотность:
1,16
pH:
4,0 (0,2 М раствор)
% Летучие по объему при 21 ° C (70 ° F):
0
Точка кипения:
Не применимый.
Точка плавления:
182C (360F) Начинает разлагаться @ 238C (460F)
Плотность пара (воздух = 1):
Информация не найдена.
Давление пара (мм рт. Ст.):
Информация не найдена.
Скорость испарения (BuAc = 1):
Информация не найдена.
10. Стабильность и реакционная способность
Стабильность:
Стабильна при обычных условиях эксплуатации и место хранения.
Опасные продукты разложения:
Двуокись углерода и окись углерода может образоваться при нагревании до разложения.
Опасно Полимеризация:
Не происходит.
Несовместимость:
Нет Обнаружены данные о несовместимости.
Условия, которых следует избегать:
Нет информации нашел.
11. Информация о токсичности
Пероральная крыса LD50 7930 мг / кг. Исследован как мутаген.
-------- \ Списки рака \ ----------------------------------- -------------------
--- Канцероген NTP ---
Известный ингредиент Ожидаемая категория IARC
------------------------------------ ----- --------- - -------------
Глицин (56-40-6) Нет Нет Нет
12.Экологическая информация
Экологическая судьба:
Информация не найдена.
Экологическая токсичность:
Информация не найдена.
13. Рекомендации по утилизации
Все, что нельзя сохранить для восстановления или переработки, следует управляются на соответствующем и утвержденном предприятии по удалению отходов. Обработка, использование или загрязнение этого продукта может изменить варианты обращения с отходами. Государственные и местные правила утилизации могут отличаться от федеральных. нормативные документы.Утилизируйте тару и неиспользованное содержимое в соответствии с федеральные, государственные и местные требования.
14. Информация о транспортировке
Не регулируется.
15. Нормативная информация
-------- \ Состояние химической инвентаризации - Часть 1 \ -------------------------------- -
Ингредиент TSCA EC Япония Австралия
----------------------------------------------- --- - --- ----- ---------
Глицин (56-40-6) Да Да Да Да
-------- \ Состояние химической инвентаризации - Часть 2 \ ---------------------------------
--Канада--
Ингредиент Корея DSL NDSL Phil.----------------------------------------------- --- - --- ---- -----
Глицин (56-40-6) Да Да Нет Да
-------- \ Федеральные, государственные и международные правила - Часть 1 \ ----------------
-SARA 302- ------ SARA 313 ------
Ингредиент RQ TPQ List Chemical Catg.
----------------------------------------- --- ----- - --- --------------
Глицин (56-40-6) Нет Нет Нет Нет
-------- \ Федеральные, государственные и международные правила - Часть 2 \ ----------------
-RCRA- -TSCA-
Ингредиент CERCLA 261.33 8 (г)
----------------------------------------- ------ --- --- ------
Глицин (56-40-6) Нет Нет Нет
Конвенция о химическом оружии: Нет TSCA 12 (b): Нет CDTA: Нет
SARA 311/312: Острый: Нет Хронический: Нет Пожар: Нет Давление: Нет
Реакционная способность: Нет (чистый / твердый)
Австралийский код Hazchem: Не выделено.
Расписание отравления: Не выделено.
WHMIS:
Настоящий паспорт безопасности материалов был подготовлен в соответствии с критериями опасности контролируемых продуктов Правила (CPR) и MSDS содержат всю информацию, требуемую СЛР.
16. Прочая информация
Рейтинги NFPA: Здоровье: 0 Воспламеняемость: 1 Реакционная способность: 0
Этикетка с предупреждением об опасности:
В составе товара производственная и личная гигиена и техника безопасности, избегайте всего ненужного воздействия химического вещества и обеспечить быстрое удаление с кожи, глаз и одежда.
Меры предосторожности на этикетке:
Нет.
Этикетка первая Aid:
Не применимо.
Использование продукта:
Лабораторный реагент.
Информация о редакции:
MSDS Раздел (ы) были изменены с момента последнего версия документа включает: 8.
Заявление об отказе от ответственности:
************************************************ **********************************************
Mallinckrodt Baker, Inc.предоставляет информацию, содержащуюся здесь в добросовестно, но не делает никаких заявлений относительно его полноты или точность. Этот документ предназначен только в качестве руководства для соответствующих осторожное обращение с материалом должным образом обученным лицом, использующим это продукт. Лица, получающие информацию, должны осуществлять свои независимые суждение при определении его пригодности для конкретной цели. MALLINCKRODT BAKER, INC. НЕ ДАЕТ НИКАКИХ ЗАЯВЛЕНИЙ ИЛИ ГАРАНТИЙ. ЯВНЫХ ИЛИ ПОДРАЗУМЕВАЕМЫХ, ВКЛЮЧАЯ БЕЗ ОГРАНИЧЕНИЙ ЛЮБЫЕ ГАРАНТИИ КОММЕРЧЕСКАЯ ЦЕННОСТЬ, ПРИГОДНОСТЬ ДЛЯ ОПРЕДЕЛЕННОЙ ЦЕЛИ В ОТНОШЕНИИ ИНФОРМАЦИЯ, УКАЗАННАЯ ЗДЕСЬ, ИЛИ ПРОДУКТ, К КОТОРЫМ СООТВЕТСТВУЕТ ИНФОРМАЦИЯ.СОГЛАСНО, MALLINCKRODT BAKER, INC. НЕ НЕСЕТ ОТВЕТСТВЕННОСТИ ЗА УБЫТКИ. РЕЗУЛЬТАТЫ ИСПОЛЬЗОВАНИЯ ДАННОЙ ИНФОРМАЦИИ.
************************************************ **********************************************
Подготовил: Здоровье и безопасность окружающей среды
Телефон: (314) 654-1600 (США)
Потребление этанола и седативный эффект изменяются у мышей, лишенных субъединицы рецептора глицина α2 — San Martin — 2020 — British Journal of Pharmacology
1 ВВЕДЕНИЕ
Прилежащее ядро (nAc) является основной входной структурой базальных ганглиев и стержневой областью в мезолимбической дофаминергической системе, опосредующей целенаправленное поведение, поведение, связанное с зависимостью, и мотивационные процессы (Di Chiara & Imperato, 1988; Russo & Nestler , 2013).Прилежащее ядро получает глутаматергическую иннервацию от медиальной префронтальной коры (ПФК), миндалины и гиппокампа, а также получает ГАМКергические и холинергические проекции из боковой перегородки. Кроме того, он посылает и принимает ГАМКергические проекции в и из вентральной тегментальной области (VTA) и получает важный дофаминергический вход из вентральной тегментальной области (Koob & Nestler, 1997; Russo & Nestler, 2013). Хорошо известно, что этанол и другие наркотики, вызывающие злоупотребление, активируют мезолимбическую дофаминергическую систему вознаграждения и повышают уровень дофамина в прилежащем ядре (Di Chiara, 1997; Jonsson, Adermark, Ericson, & Soderpalm, 2014; Soderpalm, Lido, & Ericson, 2017 ).Исследования микродиализа показали, что рецепторы глицина (GlyR) играют важную роль в повышении уровня дофамина в прилежащем ядре, продуцируемом этанолом (Molander & Soderpalm, 2005b; Soderpalm et al., 2017).
Глициновые рецепторы экспрессируются по всей ЦНС у млекопитающих и являются основными тормозными рецепторами, присутствующими в спинном мозге и стволе мозга (Legendre, 2001). Недавние исследования показали, что рецепторы глицина также экспрессируются в надспинальных областях, таких как префронтальная кора (Lu & Ye, 2011; Salling & Harrison, 2014), прилежащее ядро (Martin & Siggins, 2002; Molander & Soderpalm, 2005b; Muñoz, Yevenes, Forstera, Lovinger, & Aguayo, 2018), спинное полосатое тело (Comhair et al., 2018; Молчанова и др., 2018), ядра шва (Maguire et al., 2014) и вентральная область покрышки (Li et al., 2012; Ye et al., 2001). Экспрессия рецепторов глицина в мезолимбических областях представляет особый интерес, поскольку их присутствие может иметь отношение к полезным свойствам этанола, поскольку было продемонстрировано, что активация рецепторов глицина регулирует высвобождение дофамина и потребление этанола у грызунов (Lido, Ericson, Marston и Содерпалм, 2011; Моландер, Лоф, Стомберг, Эриксон и Содерпалм, 2005; Моландер и Содерпалм, 2005a).
Предыдущие исследования показали, что этанол усиливает функцию рецепторов глицина (Yevenes et al., 2010; Yevenes, Moraga-Cid, Peoples, Schmalzing, & Aguayo, 2008). Однако поведенческая роль глициновых рецепторов мозга в эффектах, вызванных этанолом, не была полностью изучена. Недавние данные из нескольких лабораторий показали, что рецепторы глицина важны в системе вознаграждения мозга и что α1 и α2 являются преобладающими субъединицами, экспрессируемыми в прилежащем ядре (Forstera et al., 2017; Jonsson et al., 2012). Наша лаборатория недавно охарактеризовала свойства рецепторов глицина в прилежащем ядре (Forstera et al., 2017) и определила, что шиповатые нейроны прилежащей средней части экспрессируют функциональные, чувствительные к этанолу рецепторы глицина, в первую очередь α1 и α2 субъединицы. Генетически модифицированная мышь с нокаутом (KI) для субъединицы α1 глицинового рецептора (K385 / 386A), которая имеет нарушенное взаимодействие с G-белками, продемонстрировала пониженную чувствительность к этанолу в прилежащем ядре, более высокое потребление этанола и повышенное предпочтение условного места (CPP) в этанол (Aguayo et al., 2014; Муньос и др., 2019). Поскольку известно, что α2-субъединицы глицинового рецептора регулируются этанолом (Yevenes et al., 2010), и они экспрессируются в прилежащем ядре (Forstera et al., 2017; Jonsson et al., 2012), субъединицы α2 глицинового рецептора также могут играют важную роль в регулировании мезолимбической дофаминергической схемы вознаграждения и аддиктивного поведения. Предыдущие исследования показали, что субъединицы α2 модулируют миграцию нейронов и развитие коры головного мозга (Avila et al., 2013), в то время как несколько мутаций и микроделеция в GLRA2 связаны с расстройством аутистического спектра (ASD) (Pilorge et al., 2016; Чжан, Хо, Харви, Линч и Керамидас, 2017 г.).
Целью этого исследования было оценить вклад α2-содержащих рецепторов глицина в прилежащее ядро и охарактеризовать эффекты этанола на соответствующее поведение мышей, лишенных субъединицы α2 рецептора глицина. Поскольку в предыдущих исследованиях сообщалось о половых различиях в связанном с алкоголем поведении с использованием грызунов (Becker & Koob, 2016; Caruso et al., 2018; DeFries, Wilson, Erwin, & Petersen, 1989; Rhodes et al., 2007), мы использовали самцов и самок мышей для поведенческих тестов. У мышей с нокаутом (KO) обнаружен большой дефицит экспрессии рецепторов глицина в прилежащем ядре, что продемонстрировано экспериментами с токами, вызванными глицином, и глицинергической нейротрансмиссией. Интересно, что мы обнаружили, что самцы мышей Glra2 — / Y показали увеличение потребления этанола и снижение седативного эффекта, что позволяет предположить, что мыши Glra2 KO могут быть полезной моделью для исследования расстройств, связанных с употреблением алкоголя (AUD).Тем не менее, к точным механизмам, лежащим в основе этих результатов, следует относиться с осторожностью из-за потенциальных компенсаций в других системах нейромедиаторов мозга.
2 МЕТОДА
2.1 Животные
C57BL / 6J (дикого типа [WT]) и Glra2 KO мышей (самцы: Glra2 — / Y и самки: Glra2 — / — ) в возрасте 7–12 недель. были использованы в этом исследовании. Glra2 KO мышей изначально были созданы в лабораториях Harvey and Dear путем делеции экзона 7 в гене Glra2 (Avila et al., 2013). Пары для разведения были переведены из лаборатории доктора Риго в Бельгии в Чили, где они были выведены и содержались в 12-часовом цикле свет / темнота. Glra2 KO мышей подвергали обратному скрещиванию с C57BL / 6J (IMSR Cat # JAX: 000664, RRID: IMSR_JAX: 000664) и генотипировали, как описано ранее (Avila et al., 2013). Все животные, использованные в этом исследовании, были получены от скрещиваний гемизиготных самцов ( Glra2 — / Y ) и гетерозиготных самок ( Glra2 — / + ).Уход за животными и экспериментальные процедуры были одобрены Комитетом по уходу и использованию животных Университета Консепсьона и проводились в соответствии с этическими протоколами, установленными Национальными институтами здравоохранения (NIH, Bethesda, Maryland). Об исследованиях на животных сообщается в соответствии с руководящими принципами ARRIVE (Kilkenny, Browne, Cuthill, Emerson, & Altman, 2010) и рекомендациями, сделанными British Journal of Pharmacology.
2.2 Протокол эксперимента
Все исследования были разработаны для создания групп равного размера и распределены случайным образом. Оператор и анализ данных были слепыми.
2.3 Подготовка срезов головного мозга
МышейWT и Glra2 — / Y декапитировали, как описано ранее (Forstera et al., 2017). Коронковые срезы готовили сразу после иссечения и помещения мозга в ледяной режущий раствор (в мМ: сахароза 194, NaCl 30, KCl 4.5, MgCl 2 1, NaHCO 3 26, NaH 2 PO 4 1,2 и глюкоза 10, насыщенная 95% O 2 /5% CO 2 и доведенная до pH 7,4), приклеен к охлажденному столику вибратома (Leica VT1200S, Leica Biosystems, Германия) и нарезан до толщины 300 мкм. Срезы переносили в раствор искусственного ЦСЖ (aCSF) (в мМ: NaCl 124, KCl 4,5, MgCl 2 1, NaHCO 2 26, NaH 2 PO 4 1,2, глюкоза 10 и CaCl 2 2, pH 7.4, 315–320 мОсм), насыщенный 95% O 2 /5% CO 2 при 30 ° C в течение не менее 1 часа.
2.4 Ферментативная диссоциация прилежащих нейронов
Для ферментативной диссоциации срезы мозга, содержащие прилежащее ядро, инкубировали в течение 30 минут в нормальном aCSF (насыщенном 95% O 2 /5% CO 2 ) в присутствии 0,5 мг · мл -1 проназы (Calbiochem / EDM Bioscience, Дармштадт, Германия) при 37 ° C.Прилежащее ядро отделяли от срезов, и ткань растирали через серию наконечников пипеток с уменьшающимся диаметром в 35-миллиметровой культуральной чашке в буфере для растирания (в мМ: NaCl 20, N -метил-d-глюкамин. [NMG] 130, KCl 2,5, MgCl 2 1, HEPES 10 и глюкоза 10, доведенные до pH 7,4 и 340 мОсм). Через 20 мин изолированные нейроны прикрепляли ко дну культуральной чашки и были готовы к электрофизиологическим экспериментам.
2.5 Электрофизиология
Записи были выполнены с использованием усилителя Axopatch 200B (Axon Instrument, Юнион-Сити, Калифорния) при удерживающем потенциале -60 мВ. Токи отображались и сохранялись на персональном компьютере с использованием 1322A Digidata (Axon Instruments) и анализировались с помощью Clampfit 10.1 (Axon Instruments) и MiniAnalysis 6.0 (Synaptosoft, Inc.). Пипетки-патчи с сопротивлением 4–6 МОм были приготовлены из боросиликатных микропипеток, содержащих нити (World Precision Instruments, Сарасота, Флорида, США), с использованием съемника микропипеток P-87 (Sutter Instrument, Новато, Калифорния, США) и заполнены внутренний раствор (в мМ: KCl 120, MgCl 2 4.0, BAPTA 10, Na 2 -GTP 0,5 и Na 2 -ATP 2,0, pH 7,4 и 290–310 мОсм).
2.5.1 Диссоциированные нейроны аккумбального отдела
Глицин-активированные токи были изучены в диссоциированных нейронах прилежащей кости с использованием записи целых клеток и внешнего раствора, содержащего (в мМ) NaCl 150, KCl 5,4, CaCl 2 2,0, MgCl 2 1,0, глюкозу 10 и HEPES 10 ( pH 7,4, 315–320 мОсм). Амплитуду глицинового тока измеряли с помощью короткого импульса (1-2 с) различных концентраций глицина.Мы использовали набор внешних трубок (внутренний диаметр 200 мкм), помещенных в пределах 50 мкм от нейрона, и растворы, содержащие лиганды, непрерывно вытекали из трубок под действием силы тяжести.
2.5.2 Запись срезов мозга
Корональные срезы головного мозга (300 мкм), содержащие область прилежащего ядра, использовали для электрофизиологических записей. Глицинергические миниатюрные ингибирующие постсинаптические токи (mIPSC) были фармакологически изолированы с помощью ванны с использованием 6-циано-7-нитрохиноксалин-2,3-диона (CNQX, 10 мкМ) в качестве антагониста рецептора AMPA; бикукуллин (10 мкМ) как антагонист ГАМК А ; мекамиламин (10 мкМ) как антагонист никотиновых рецепторов ACh и тетродотоксин (ТТХ, 500 нМ).Анализ частоты (Гц), константы затухания (мс) и амплитуды (pA) использовался для сравнения мышей WT и Glra2 — / Y . Фаза затухания описывалась одной экспоненциальной кривой и составляла от 10% до 90% ее амплитуды.
2.6 Количественная ПЦР в реальном времени
Для анализа относительной экспрессии генов мышей WT и Glra2 — / Y мышей в возрасте 9–16 недель анестезировали изофлураном и умерщвляли декапитацией.По возможности ткани каждого животного использовались для нескольких экспериментов. Ткань Nucleus accumbens мышей WT собирали и хранили в течение 5–14 дней при -80 ° C в TRIzol (Ambion, Life Technologies, UK) перед дальнейшей обработкой. Тотальную РНК выделяли из образцов с использованием реагента TRIzol и хлороформа (Дармштадт, Германия) с последующим расщеплением ДНКазой I (Thermo Scientific, Уолтем, Массачусетс, США) в течение 30 мин. кДНК получали из 0,5 мкг тотальной РНК с помощью набора для синтеза кДНК Affinity Script qPCR (Agilent Technologies, Санта-Клара, Калифорния, США), включая получение отрицательных контролей без образцов РНК (H 2 O).ПЦР выполняли в двух экземплярах с использованием 1 мкл кДНК в течение 40 циклов с температурой отжига 57 ° C в циклере Strategene Mx3005P (Agilent Technologies) для количественной ПЦР в реальном времени (qPCR) ( n = 5–6 в WT; n = 6 дюймов Glra2 — / Y ) с использованием Brilliant II SYBR QPCR Master Mix (Agilent Technologies), включая кривые плавления для контроля специфичности qPCR. Данные qPCR были собраны и проанализированы с помощью MxPro (Agilent Technologies), и был использован метод δ – δ Ct.18S использовали в качестве контрольного гена, а пороговые циклы экспрессии рецептора глицина были нормализованы до 18S. Праймеры, используемые для qPCR в реальном времени, показаны в таблице S1. Праймеры, используемые для обнаружения мРНК субъединицы α2, гибридизуются с экзоном 7, который соответствует удаленной области гена Glra2 в линии мышей KO.
2,7 Вестерн-блоттинг
Гомогенаты тканей из прилежащего ядра (50 мкг) после обработки лизисом (10-мМ трис-HCl, pH 7.4, 0,25-М сахарозу, 10-мМ N -этилмалеимид [NEM] и смесь ингибиторов протеаз 1X) загружали в 10% SDS-PAGE и помещали в камеру для электрофореза. Затем белки наносили на нитроцеллюлозные мембраны, блокировали 5% -ным молоком в трис-буферном солевом растворе (TBS) с 0,1% Tween 20 в течение 1 ч и инкубировали с первичными антителами против пан-рецептора α-глицина (1: 500, кроличий моноклональный IgG). ; № по каталогу 146008, Synaptic Systems, RRID: AB_2636914), антитело против рецептора глицина β (1: 200, кроличий поликлональный IgG; номер по каталогу AGR-014, Alomone, RRID: AB_2340973) и антитело против Gβ (1: 1000 , кроличий поликлональный IgG; № по каталогу sc-378, Santa Cruz Biotechnology, RRID: AB_631542) в течение ночи при 4 ° C.После стадий промывки мембраны инкубировали с вторичными антителами против кролика, конъюгированными с HRP (1: 5,000, поликлональные козьи антитела против IgG-HRP кролика, Cat # sc-2004, Santa Cruz Biotechnology, RRID: AB_631746). Иммунореактивность белков определяли с использованием системы обнаружения вестерн-блоттинга ECL Plus (PerkinElmer, Бостон, Массачусетс). Относительную экспрессию белка нормализовали с использованием экспрессии субъединицы Gβ. Используемые иммуно-связанные процедуры соответствуют рекомендациям British Journal of Pharmacology (Alexander et al., 2018).
2.8 Поведенческие исследования
В этом исследовании использовали мышей-самцов ( Glra2 — / Y ) и самок ( Glra2 — / — ), если не указано иное. Мышам позволяли акклиматизироваться в экспериментальной комнате в течение не менее 1 часа перед поведенческими анализами. Этанол разбавляли 0,9% физиологическим раствором (20% об. / Об.) И вводили внутрибрюшинными инъекциями в дозах, скорректированных в соответствии с введенными объемами.
2.8.1 Анализ открытого поля
Мышей тестировали на базальную двигательную активность в новом открытом поле с использованием системы видеонаблюдения (ANY-maze, Stoelting Co.). Мышам позволяли свободно исследовать тестовую область размером 50 × 50 см в течение 25 мин. Было проанализировано общее пройденное расстояние и время нахождения в центре.
2.8.2 Роторный ускоритель
Показатели базальной моторики были проверены с помощью теста с ускоряющим вращающимся стержнем.Мышей помещали на невращающийся стержень (IITC, Life Science). Испытание началось, когда штанга начала разгоняться с 4 до 40 об / мин за 120 с, и была зафиксирована задержка до падения. Каждую мышь тестировали 5 раз в 1-й день и 10 раз во 2-й день.
2.8.3 Ротор с фиксированной скоростью
Мышей тестировали на чувствительность и толерантность к моторным атаксическим эффектам этанола с использованием теста с фиксированной скоростью вращения стержня (IITC, Life Science). Мышей обучали (скорость вращения стержня 5 об / мин) в течение 3 дней, и обучение считали завершенным, когда мыши могли оставаться на вращающемся стержне в течение 120 с.Мышам вводили этанол (2,0 г · кг -1 внутрибрюшинно), и каждые 15 минут после инъекции мышей помещали обратно на вращающуюся головку и измеряли латентность падения до тех пор, пока мыши не смогли полностью оставаться на вращающейся головке. 120 с.
2.8.4 Потеря рефлекса выпрямления
Мышей тестировали на седативный эффект этанола с использованием 3,5 г · кг -1 i.p. Мышам вводили этанол, и когда мыши становились атаксичными, их помещали в положение лежа на спине в V-образные пластиковые лотки до тех пор, пока они не смогли трижды выпрямиться в течение 30 секунд.Потеря рефлекса выпрямления (LORR) определялась как время от помещения в положение лежа на спине до восстановления рефлекса выпрямления. Эти эксперименты проводились в Чили и Бельгии. Хотя мы обнаружили аналогичные результаты в отношении продолжительности потери рефлекса выпрямления между двумя лабораториями, начало потери рефлекса выпрямления было различным. Мы связали эти различия со скоростью внутрибрюшинной инъекции, диетой мышей и / или фоновым шумом в обоих местах. Тем не менее, сходство результатов по продолжительности потери рефлекса выпрямления очень важно, потому что это наиболее широко используемый показатель седации / интоксикации этанолом в нашей и других лабораториях (Aguayo et al., 2014; Бледнов и др., 2015; Бледнов, Бенавидес, Гоманикс и Харрис, 2012 г.).
2.8.5 Пить в темноте (DID)
Этот тест на употребление алкоголя с ограниченным доступом производит значительные уровни этанола в крови (Rhodes, Best, Belknap, Finn, & Crabbe, 2005). Мышей переводили в индивидуальные клетки и давали им акклиматизироваться не менее 1 недели. Через два часа после выключения света бутылки с водой были заменены бутылками, содержащими 15% об. / Об. Раствора этанола, либо на 2 часа в течение первых 3 дней, либо на 4 часа на четвертый день.Бутылки с этанолом взвешивали перед размещением и после извлечения из клеток каждый день. Количество израсходованного этанола рассчитывали как -1 г · кг массы тела за 2 или 4 часа соответственно.
2.8.6 Концентрация этанола в крови
Образцы крови из лицевой вены от WT и Glra2 — / Y (или Glra2 — / — ) были собраны через 10 минут в первый и четвертый день питья в темноте.Образцы крови центрифугировали (10000 об / мин × 10 мин) и концентрацию этанола в сыворотке определяли с помощью анализатора Analox AM1 (Люненбург, Массачусетс).
2.8.7 Условное предпочтение места
Условное предпочтение места было выполнено, как описано ранее (Muñoz et al., 2019). Эта парадигма основана на Павловском обусловливающем поведении. Шесть идентичных акриловых ящиков (33 × 27 × 20 см) отдельно помещались в вентилируемые, свето- и звукопоглощающие камеры.Вкратце, исследование кондиционирования места включало сеанс предварительной подготовки, восемь сеансов кондиционирования и сеанс посткондиционирования. Для сеанса предварительного кондиционирования мыши получали инъекцию физиологического раствора непосредственно перед тем, как их поместили в бокс, где оба отсека были доступны на 15 мин. Было проведено восемь сеансов кондиционирования, и мыши получали инъекцию либо этанола 2,0 г · кг -1 (Cs +), либо физиологического раствора (Cs-) через день, и их движение регистрировалось в течение 10 минут. В отсеках имелся разный размер решетки пола: 0.Сетка 6 см для положительного стимула (Cs +) и сетка 0,1 см для отрицательного стимула (Cs−). Наконец, для посткондиционирования мыши получали инъекцию физиологического раствора перед тем, как их помещали в центр бокса без разделения для проверки активности в 30-минутном сеансе предпочтений. Предпочтение записи видео анализировалось с помощью системы видеонаблюдения (программное обеспечение ANY-maze, Stoelting Co.).
2,9 Реагенты
Бикукуллин и CNQX были приобретены у Tocris (Бристоль, Великобритания).Глицин и стрихнин (STN) были получены от Sigma-Aldrich (США). ТТХ был приобретен в Alomone labs (Иерусалим, Израиль). Этанол был приобретен в Merck Millipore (США).
2.10 Анализ данных
Результаты выражены в виде среднего значения ± стандартная ошибка, и статистический анализ был выполнен с использованием непарного теста Стьюдента t или двухфакторного дисперсионного анализа с повторными измерениями для исследований, в которых размер каждой группы составлял не менее n = 5. Размер группы в этом исследование представляет собой независимые ценности.Статистически значимыми считались различия с P <0,05. После ANOVA апостериорный тест Бонферрони запускали, только если F достигал необходимого уровня статистической значимости ( P <0,05) и не было значительной неоднородности дисперсии. Как и в предыдущих исследованиях (Aguayo et al., 2014; Muñoz et al., 2019), чтобы получить статистическую мощность выше 95% ( α, = 0,05, мощность = 0,95), чтобы определить наличие статистически значимых различий ( P <0.05), мы использовали выборку из 6–8 измерений для экспериментальной группы. Для изучения поведения мы рассматривали не менее 10–12 животных в группе; однако были небольшие различия в размере групп из-за ненадежных внутрибрюшинных инъекций или проблем с бутылками. Программное обеспечение OriginPro 9.0 (Microcal Origin, RRID: SCR_002815, Northampton, MA, USA) использовалось для всех статистических анализов. Данные и статистический анализ соответствуют рекомендациям British Journal of Pharmacology по экспериментальному дизайну и анализу в фармакологии (Curtis et al., 2018).
2.11 Номенклатура мишеней и лигандов
Ключевые белковые мишени и лиганды в этой статье имеют гиперссылки на соответствующие записи в http://www.guidetopharmacology.org, общем портале данных из IUPHAR / BPS Guide to PHARMACOLOGY (Harding et al., 2018), и постоянно Архивировано в Кратком руководстве по ФАРМАКОЛОГИИ 2019/20 (Александр и др., 2019).
3 РЕЗУЛЬТАТЫ
3.1 Наличие α2-содержащих рецепторов глицина в прилежащем ядре
Предыдущие исследования показали, что прилежащее ядро экспрессирует α и β субъединицы глицинового рецептора (Jonsson et al., 2012; Muñoz et al., 2019). Анализ относительной экспрессии субъединицы глицинового рецептора в прилежащем ядре с помощью ПЦР в реальном времени выявил высокие уровни субъединиц α2 и β у мышей WT, в то время как у мышей Glra2 — / Y экспрессия α2 полностью исчезла. , как и ожидалось (рис. 1а).Интересно, что уровни экспрессии α1 и α3 были увеличены, но были значимыми только для α3 у мышей Glra2 — / Y по сравнению с WT, в то время как уровни экспрессии β, с другой стороны, неожиданно значительно снизились. мыши, лишенные субъединицы α2. Чтобы изучить уровни экспрессии белка, мы провели эксперименты вестерн-блоттинга в лизатах тканей из прилежащего ядра, выделенных из срезов мозга, с использованием антител к субъединицам α и β. Предварительные результаты показали, что субъединица α2 составляет важный компонент рецепторов глицина, экспрессируемых в нейронах прилежащей зоны у мышей WT, поскольку экспрессия общих субъединиц α у мышей, лишенных субъединиц α2, была значительно снижена, что отражено в денситометрическом анализе (рис. 1b).Присутствие субъединицы β, белка, важного для закрепления рецепторов глицина на синаптических сайтах (Grudzinska et al., 2005), также было проанализировано на мышах Glra2 — / Y с помощью вестерн-блоттинга. Неожиданно результаты не выявили значительных изменений в экспрессии субъединиц β у мышей Glra2 — / Y по сравнению с мышами WT (рис. 1c). Хотя этот результат противоречит анализу КПЦР транскрипта субъединицы β, это может быть связано с различиями в стабильности мРНК по сравнению с белком.
Наличие субъединиц α и β глицинового рецептора (GlyR) в прилежащем ядре (nAc) мышей WT и Glra2 — / Y . (а) Количественная ПЦР в реальном времени субъединиц GlyR в nAc мышей дикого типа (WT) и Glra2 — / Y мышей. (b, c) Вестерн-блоттинг nAc от мышей WT и Glra2 — / Y для определения α- и β-субъединиц GlyR. Графики показывают низкие уровни α-субъединиц у мышей Glra2 — / Y по сравнению с мышами WT ( n = 3 WT и n = 3 Glra2 — / Y ) и т.п. уровни субъединицы β у обеих мышей ( n = 4 WT и n = 4 Glra2 — / Y ).Непарный Студенческий т -тест. Данные представляют собой среднее значение ± стандартная ошибка среднего. * P <0,05
3.2 Потеря активируемых глицином токов в нейронах прилежащей области у мышей
Glra2 — / YНедавние результаты нашей лаборатории показали, что в прилежащих нейронах присутствуют функциональные рецепторы глицина, чувствительные к этанолу (Forstera et al., 2017), и что α1 и α2 являются преобладающими субъединицами, экспрессируемыми в этой области (Forstera et al., 2017; Jonsson et al., 2012). Используя электрофизиологические записи в диссоциированных нейронах (возрастом до 8 недель), мы обнаружили, что только 17% нейронов Glra2 — / Y (4 из 24 прилежащих нейронов) имели амплитуды более 30 пА с использованием насыщающая концентрация глицина (рис. 2b). Эти нейроны показали очень небольшие токи, вызванные глицином (~ 100 пА на 1 мМ глицина в этих четырех клетках) (рис. 2d), по сравнению с мышами WT (~ 500 пА при 1 мМ глицина) (рис. 2a).Анализ плотности тока показал, что мыши Glra2 — / Y имели значительно меньшие значения (1,7 ± 0,7 пФ · pA -1 ) по сравнению с мышами WT (39,5 ± 6,3 пФ · pA -1 ; 2в). Интересно, что плотность тока ответов ГАМК также была значительно снижена у мышей Glra2 — / Y (49,2 ± 5,1 пФ · pA −1 по сравнению с 73,3 ± 7,7 пФ · pA −1 для WT, Рисунок 2в). Кривая концентрация глицина – ответ для четырех нейронов мышей Glra2 — / Y показала ЕС 50 55 ± 8 мкМ, в то время как мыши WT показали ЕС 50 64 ± 5 мкМ (рис. 2г).Для количественной оценки потенцирования этанолом использовалась концентрация глицина, которая активирует 10-15% максимального тока (EC 10 ) (Yevenes et al., 2008). Из-за малой амплитуды вызванного глицином тока, полученного в этих Glra2 — / Y нейронах, было невозможно оценить чувствительность к этанолу, сделав вывод, что акумбальные рецепторы глицина в основном отсутствуют у мышей KO. Это было по меньшей мере удивительно, потому что мы предсказали только снижение тока, активируемого глицином, из-за присутствия субъединиц α1 и α3 (см. Раздел 4).
Глициновый ответ прилежащих нейронов от мышей Glra2 — / Y . (а) Типичные текущие следы, индуцированные насыщающей концентрацией агониста в диссоциированных нейронах из прилежащего ядра (nAc) мышей WT и Glra2 — / Y . (b) Только 4 из 24 нейронов прилежащей кости (17%) мышей Glra2 — / Y имели вызванный глицином ток более 30 пА.(c) График показывает плотность тока в аккумбальных нейронах мышей WT и Glra2 — / Y . (d) Репрезентативные следы вызванных глицином токов от отвечающих нейронов (1–1000 мкМ) у мышей Glra2 — / Y . На графике показана кривая «концентрация глицина-ответ» в прилежащих нейронах мышей WT (черные квадраты) и Glra2 — / Y (красные квадраты). Непарный Студенческий т -тест. Данные представляют собой среднее значение ± стандартная ошибка среднего.* P <0,05
3.3 Субъединица α2 глицинового рецептора важна для глицинергических синаптических токов
Недавно опубликованная статья показала наличие глицинергических синаптических токов в прилежащих нейронах (Muñoz et al., 2018). Это было подтверждено вестерн-блоттингом и иммуногистохимическим анализом, который показал присутствие субъединиц α и β и GlyT2, пресинаптического переносчика глицина (Bradaia, Schlichter, & Trouslard, 2004).В настоящем исследовании мы хотели оценить вклад субъединиц α2 глицинового рецептора в глицинергические входы прилежащего ядра. Используя смесь ингибиторов (ТТХ, бикукуллин, CNQX и мекамиламин), мы выделили глицинергические синаптические токи в срезах, содержащих прилежащее ядро. Мы обнаружили, что 64% от общего числа нейронов мышей WT обладали глицинергической активностью, тогда как только 38% от общего числа нейронов мышей Glra2 — / Y имели глицинергические синаптические токи (рис. 3а).На рисунке 3b показаны репрезентативные следы общих миниатюрных постсинаптических токов (мПСК), зарегистрированных в прилежащем ядре мышей Glra2 — / Y в отсутствие ингибиторов. После фармакологического выделения миниатюрных глицинергических постсинаптических токов (глицинергических mIPSC) мы обнаружили небольшое количество синаптических событий. Полная блокада этих миниатюрных событий в присутствии стрихнина свидетельствовала о глицинергических синаптических токах (рис. 3b). Средние трассы этих событий показаны на рисунке 3c.Анализ синаптических параметров показал значительное снижение частоты общих mPSC у мышей Glra2 — / Y (0,81 ± 0,24 Гц) по сравнению с WT (2,50 ± 0,28 Гц) и значительное снижение частоты фармакологически изолированных глицинергических mIPSC. у мышей Glra2 — / Y (0,68 ± 0,09 Гц у мышей WT против 0,17 ± 0,06 Гц у мышей Glra2 — / Y , рис. 3d). При анализе амплитуды изменений между генотипами для общих мПСК не обнаружено (15.9 ± 1,3 пА у мышей WT по сравнению с 15,9 ± 1,7 пА у мышей Glra2 — / Y ) или глицинергических mIPSC (8,4 ± 0,7 пА у мышей WT по сравнению с 10,2 ± 0,8 пА у Glra2 — / Y мышей, рис. 3д). Наконец, анализ константы распада показал, что общие mPSC показали сходные значения для обоих генотипов (11,2 ± 0,9 мс у мышей WT против 12,3 ± 2,3 мс у Glra2 — / Y mic). Однако наблюдалось увеличение, хотя и незначительное, в константе распада глицинергических mIPSCs у Glra2 — / Y мышей из 12.5 ± 2,7 мс по сравнению с 7,7 ± 0,9 мс у мышей WT (рис. 3f). Поскольку прилежащие глицинергические токи у мышей WT были нечувствительны к этанолу (Muñoz et al., 2018), мы не изучали влияние этанола на глицинергические токи прилежащего отдела у мышей Glra2 — / Y .
Синаптическая активность в аккумбальных нейронах мышей WT и Glra2 — / Y . (а) Процент нейронов, которые имеют глицинергические синаптические токи от прилежащих нейронов мышей WT и Glra2 — / Y .(b) Репрезентативные следы синаптического тока от мышей Glra2 — / Y для общего миниатюрного постсинаптического тока (mPSC), фармакологически изолированного глицинергического mIPSC и глицинергического mIPSC, блокированного стрихнином (STN). (c) Средние значения для mPSC (черный), глицинергического mIPSC (синий) и STN (зеленый). (d – f) На графиках обобщены синаптические параметры: (d) частота, (e) амплитуда и (f) константа распада от mPSCs и глицинергических mIPSCs в прилежащих нейронах от WT и Glra2 — / Y мышей ( n = 14 WT и n = 13 Glra2 — / Y мышей).Непарный тест Студента t- . Данные представляют собой среднее значение ± стандартная ошибка среднего. * P <0,05
3,4
Glra2 KO-мыши показали нормальную двигательную активность, но с нарушением обучения двигательным навыкамПоскольку Glra2 расположен на X-хромосоме, все взрослые мужчины, использованные в поведенческом исследовании, были гемизиготными ( Glra2 — / Y ), в то время как женщины были гомозиготными ( Glra2 — / — ) для делеция экзона 7.Подобно предыдущим исследованиям (Pilorge et al., 2016), эта генетически модифицированная мышь не демонстрировала поведения сцепления лап, когда мышь поднималась за хвост (рисунок S1A), что указывает на отсутствие изменений мышечного тонуса и двигательных рефлексов. Кроме того, с использованием качественного теста на захват, мыши Glra2 — / Y не показали серьезных изменений мышечной силы (рисунок S1B). Мыши Glra2 — / Y показали нормальный мозг и общий вес по сравнению с мышами WT (фигура S1C, D).
Базальную двигательную активность изучали с помощью теста открытого поля как на мышах WT, так и на мышах Glra2 KO. Первоначальный анализ пола показал, что не было значительных различий в общем расстоянии, пройденном между самцами и самками у мышей WT и у мышей Glra2 KO. Поэтому мы объединили данные обоих полов в этом анализе. Анализ общего пройденного расстояния не показал различий между мышами WT и мышами Glra2 KO (рис. 4a, b).Также не было различий между генотипами, когда мы анализировали время, проведенное в центре, индикатор тревожно-подобного поведения (рис. 4c).
Базальная двигательная активность мышей с нокаутом Glra2 ( Glra2 KO). (а) Было исследовано расстояние, пройденное в открытом поле для проверки двигательной активности. Репрезентативные траектории мыши WT и мыши с нокаутом Glra2 в течение первых 5 мин в тесте в открытом поле.(b) График показывает общее расстояние, пройденное за 20 минут, количественно каждые 5 минут. (c) Не было обнаружено различий во времени, проведенном мышами в центре. В этом тесте использовали как самцов, так и самок ( n = 29 WT и n = 25 Glra2 KO мышей). Двусторонний дисперсионный анализ с повторными измерениями. Данные являются средними ± SEM
.Когда мы провели анализ с помощью ускоряющего вращающегося стержня, мы обнаружили, что латентность падения была значительно увеличена у самок WT по сравнению с самцами мышей WT (т.е., для испытания 15: 70 ± 4 против 57 ± 4 с). Поэтому мы решили разделить данные по полу. Интересно, что в этом эксперименте латентность падения была значительно увеличена у самцов мышей Glra2 — / Y по сравнению с самцами мышей WT (рис. 5a). Кроме того, испытание имело значительный эффект. Аналогичным образом, анализ латентности падения у самок мышей также продемонстрировал, что мыши Glra2 — / — работали значительно лучше, чем мыши дикого типа (рисунок S2A).Также наблюдался значительный эффект испытания, указывающий на разницу в двигательных навыках между полами и генотипами.
Эффект высоких доз этанола у мышей-самцов с нокаутом Glra2 ( Glra2 — / Y ). (а) Мышей тестировали на двигательные навыки с использованием ускоряющего вращающегося стержня, который состоял из нескольких испытаний в течение 2-дневного периода. График выявил значительную разницу в производительности: мыши Glra2 — / Y имели увеличенную задержку падения по сравнению с мышами WT ( n = 12 WT и n = 12 Glra2 — / Y мышей).(b) Атаксический эффект 2,0 г · кг -1 этанола на тесте вращающегося стержня с фиксированной скоростью у самцов мышей WT и Glra2 — / Y мышей. Мышей обучали в течение 3 дней до максимального времени 120 с при 8 об / мин. На 4 день мышам внутрибрюшинно вводили этанол и тестировали каждые 15 минут после инъекции. Не было обнаружено значительных различий между генотипами ( n = 12 WT и n = 11 Glra2 — / Y ). Двусторонний дисперсионный анализ с повторными измерениями и апостериорный тест Бонферрони.(c) Потеря выпрямляющего рефлекса (LORR) использовалась как показатель депрессии ЦНС. Мышам внутрибрюшинно вводили 3,5 г · кг -1 этанола и измеряли латентный период до LORR. Мыши Glra2 — / Y теряли свою рефлекторную способность быстрее, чем мыши WT. (d) Способность восстанавливать рефлекс выпрямления измерялась как продолжительность LORR, где мыши Glra2 — / Y восстанавливались быстрее, чем мыши WT ( n = 12 WT и n = 10 Glra2 ). — / Y ).Непарный студенческий т -ест. Данные представляют собой среднее значение ± стандартная ошибка среднего. * P <0,05
3,5 Снижение вызванной этанолом седации у мышей, лишенных субъединицы α2 рецептора глицина
Мышей также тестировали с использованием высоких доз этанола для изучения моторных атаксических эффектов и седативного эффекта. Используя тест с фиксированной скоростью вращающегося стержня, мы проверяли двигательную активность каждые 15 минут после внутрибрюшинной инъекции 2,0 г · кг -1 этанола. Поскольку мы обнаружили, что самки WT выздоравливали значительно быстрее, чем самцы мышей WT (т.е.е. минута 60: 118 ± 2 против 80 ± 8 с; минута 75: 118 ± 2 против 102 ± 6 с), данные также были разделены по полу. После инъекции этанола мы не обнаружили различий в восстановлении после атаксии между генотипами у самцов (рисунок 5b) и самок мышей (рисунок S2B). Важно отметить, что оба генотипа смогли проявить себя во время тренировочных испытаний перед инъекцией.
Интересно, что когда мы исследовали самцов мышей Glra2 — / Y на потерю рефлекса выпрямления в присутствии седативной дозы этанола (3.5 г · кг -1 этанола ip), мы обнаружили значительное снижение начала потери рефлекса выпрямления (2,3 ± 0,2 мин для WT; 1,7 ± 0,1 мин для Glra2 — / Y ) и продолжительность потери рефлекса выпрямления (39 ± 3 мин для WT; 27 ± 3 мин для Glra2 — / Y ) (рис. 5в, г). Тот же анализ был проведен в лаборатории нашего сотрудника в Бельгии, где также было обнаружено, что продолжительность потери рефлекса выпрямления была уменьшена у мышей Glra2 – / Y по сравнению с мышами дикого типа (рис. S3B).Однако начало потери рефлекса выпрямления было увеличено у мышей Glra2 — / Y по сравнению с мышами дикого типа (рис. S3A), что, вероятно, могло быть связано с меньшей скоростью внутрибрюшинной инъекции, диетой мышей и / или фоновый шум. Анализ на самках мышей показал, что потеря рефлекса выпрямления не изменилась у мышей Glra2 — / — (рисунок S2C, D). Интересно отметить, что продолжительность потери рефлекса выпрямления значительно различалась между полами у мышей WT (39 ± 3 мин для самцов vs.28 ± 2 мин для самок). Чтобы подтвердить, что эти различия не связаны с фармакокинетикой этанола, концентрации этанола в крови измеряли через 30 минут после внутрибрюшинной инъекции этанола (3,5 г · кг -1 ). Концентрация этанола в крови у самцов мышей составляла 394 ± 14 и 429 ± 15 мг · дл -1 для мышей WT и Glra2 — / Y соответственно, тогда как у самок мышей значения составляли 419 ± 24 и 435 ± 8 мг · дл -1 для мышей WT и Glra2 — / — соответственно.Таким образом, у женщин потеря рефлекса выпрямления была короче, чем у мужчин с аналогичными значениями концентрации этанола в крови.
3,6
Glra2 — / Y Мыши потребляли больше этанола при первом воздействии по сравнению с мышами дикого типаНесколько отчетов продемонстрировали, что рецепторы глицина, присутствующие в системе вознаграждения, важны для потребления и предпочтения этанола (Lido et al., 2011; Molander et al., 2005). Точно так же на повышение уровней дофамина, продуцируемого этанолом в прилежащем ядре, влияет активация рецептора глицина (Molander & Soderpalm, 2005a).Мы использовали парадигму употребления алкоголя в темноте, как описано ранее (Rhodes et al., 2005), для оценки поведения, связанного с потреблением. Первоначальный анализ показал, что существует значительная разница в потреблении этанола между самцами и самками мышей WT. Поэтому мы разбиваем данные по полу для дальнейшего анализа. Наши результаты показали, что самцы дикого типа начали постепенно переедать в течение 4 дней анализа. С другой стороны, мыши Glra2 — / Y потребляли значительно больший этанол (уровни переедания) по сравнению с самцами мышей WT ( P <0.05) уже в дни 1 и 2 (день 1: 2,3 ± 0,2 г · кг −1 в WT против 3,0 ± 0,2 г · кг −1 в Glra2 — / Y ; день 2 : 2,4 ± 0,2 г · кг -1 у мышей WT по сравнению с 3,5 ± 0,3 у Glra2 — / Y , рисунок 6a). Анализ концентрации этанола в крови показал, что мыши Glra2 — / Y имели значительно более высокие уровни этанола в крови в первый день потребления по сравнению с мышами WT (62 ± 10 мг · дл -1 в WT. против.106 ± 18 мг · дл -1 дюйм Glra2 — / Y , рис. 6b). Интересно, что уровни у мышей Glra2, — / Y были выше предела запоя, установленного Национальным институтом злоупотребления алкоголем и алкоголизма. Анализ концентрации этанола в крови на 4 день, через 4 часа потребления, не выявил значимых различий между генотипами (149 ± 24 мг · дл -1 для WT против 116 ± 26 мг · дл -1 для Glra2. — / Y , рисунок 6c).Когда эксперимент проводился на самках, мы обнаружили аналогичную картину потребления у самок мышей Glra2 — / — по сравнению с самцами мышей Glra2 — / Y (рисунок 6d), но различия были не является статистически значимым при сравнении самок мышей WT и Glra2 — / — мышей. Как сообщалось ранее (Rhodes et al., 2007), самки мышей WT потребляли больше этанола, чем самцы мышей WT, при этом значения достигали 3.8 ± 0,3 г · кг -1 для женщин и 2,3 ± 0,2 г · кг -1 для мужчин в первый день потребления (рис. 6d). Анализ концентрации этанола в крови на 4 день показал, что самки мышей не показали различий между генотипами (178 ± 14 мг · дл -1 для WT против 166 ± 16 мг · дл -1 для Glra2 — / — , рисунок 6д).
Самцы мышей с нокаутом Glra2 потребляют большее количество этанола при питье в темноте.(а) График суммирует результаты теста питья в темноте (DID) у самцов мышей WT и мышей Glra2 — / Y . Во время темной фазы дня мыши могли пить 15% об. Раствор этанола в течение 2 часов в течение первых 3 дней. На четвертый день мышам давали пить в течение 4 часов. Самцы мышей Glra2 — / Y потребляли больше этанола в дни 1 и 2 по сравнению с мышами WT (измерено в граммах этанола на килограмм веса [г · кг -1 ]; n = 8 WT и n = 9 Glra2 — / Y ).Двусторонний дисперсионный анализ с повторными измерениями и апостериорный тест Бонферрони. (b) График показывает, что после первого дня потребления мыши Glra2 — / Y имели более высокие концентрации этанола в крови (BEC), чем мыши WT. (c) БЭК на 4-й день показал аналогичные уровни для этанола. Непарный Студенческий т -тест. (d) На графике суммированы результаты теста DID на самках мышей WT и Glra2 — / — ( n = 8 WT и n = 10 Glra2 — / — ).(e) Анализ BEC показывает отсутствие различий в уровнях этанола между генотипами. Непарный Студенческий т -тест. Данные представляют собой среднее значение ± стандартная ошибка среднего. н.с. P > 0,05, * P <0,05
3,7 Снижение предпочтения мест, обусловленных этанолом, у мышей
Glra2 — / YНаконец, мы выполнили условный тест предпочтения места для оценки предпочтения этанола мышей WT и Glra2 — / Y .Этот тест представляет собой форму Павловского кондиционирования, используемую для измерения поощрительных и мотивационных эффектов злоупотребления наркотиками (Cunningham, Gremel, & Groblewski, 2006). Ранее сообщалось, что мыши KI для субъединицы рецептора глицина α1 показали повышенное предпочтение стороны этанола по сравнению с мышами WT, демонстрируя важность рецепторов глицина в регуляции поведения, связанного с поиском алкоголя (Muñoz et al., 2019). Мыши WT и Glra2 — / Y , использованные в тесте предпочтения кондиционированного места, не показали предпочтения ни одной из сторон клетки во время 15-минутного сеанса предварительного кондиционирования (рис. 7a, c).После восьми сеансов кондиционирования мыши WT имели значительное предпочтение в отношении инъекции этанола (Cs +) в течение 30-минутного сеанса после кондиционирования (35 ± 2 против 25 ± 2 с · мин -1 , рис. 7b, d). . Интересно, что у мышей Glra2 — / Y после восьми сеансов кондиционирования не было обнаружено предпочтения этанола (31 ± 3 против 29 ± 3 с · мин -1 , фиг. 7b, d). Эти результаты показывают, что мыши Glra2, — / Y утратили предпочтение этанола по сравнению с дикими животными.
Отсутствие изменений в предпочтении мест, обусловленных этанолом, у самцов мышей Glra2 — / Y . (а) Репрезентативные следы траектории, полученные в 15-минутном сеансе предварительной подготовки, полученные от мышей WT (слева) и Glra2 — / Y (справа). (b) Репрезентативные следы траектории в 30-минутном сеансе посткондиционирования, полученные от мышей WT (слева) и Glra2 — / Y (справа).(c) График суммирует предпочтение места для предварительного кондиционирования, полученное у мышей WT и Glra2 — / Y , и не показывает различий во времени, проведенном на стороне этанола (Cs +) по сравнению со стороной носителя (Cs− ). (d) График суммирует предпочтение места после кондиционирования после восьми кондиционирования мышей WT и Glra2 — / Y . Результаты показали, что мыши WT проводили больше времени на стороне этанола (Cs +), чем на стороне носителя (Cs−), в то время как мыши Glra2 — / Y не показали предпочтения ни одной из сторон ( n = 12 WT и n = 12 Glra2 — / Y ).Двусторонний дисперсионный анализ с повторными измерениями и апостериорный тест Бонферрони. Данные представляют собой среднее значение ± стандартная ошибка среднего. * P <0,05
4 ОБСУЖДЕНИЕ
Функция рецепторов глицина в надспинальных областях привлекает все большее внимание к поведению, связанному с обучением, основанным на вознаграждении, и потреблением этанола (Burgos, Munoz, Guzman, & Aguayo, 2015; Soderpalm et al., 2017). Частично это связано с тем, что в настоящее время существуют генетически модифицированные модели животных, которые позволяют изучать поведение при отсутствии значительных неврологических изменений, которые могут возникнуть при модификации критического белка мембраны мозга, такого как лиганд-зависимые ионные каналы (Aguayo и другие., 2014; Бледнов и др., 2003; Бледнов и др., 2012). Предыдущие исследования показали, что эти рецепторы не присутствуют в полосатом теле (Malosio, Marqueze-Pouey, Kuhse, & Betz, 1991). Однако недавние исследования установили, что они широко экспрессируются в большинстве областей мозга (Forstera et al., 2017; Jonsson et al., 2012). Несмотря на это, конкретные подтипы рецепторов глицина, экспрессируемые в этих областях, остаются в значительной степени неизвестными. Предыдущие исследования с использованием электрофизиологии, количественной ПЦР и иммуногистохимии показали присутствие субъединиц α1 и α2 рецепторов глицина в прилежащем ядре (Forstera et al., 2017; Муньос и др., 2018; Муньос и др., 2019). Исследования на мышах knock in (KI) показали важность α1-субъединицы в потенцировании рецептора глицина при клинически значимых концентрациях этанола, а также в потреблении и предпочтении этанола (Aguayo et al., 2014; Muñoz et al., 2019). Однако возможный вклад субъединиц α2 рецептора глицина в эффекты, связанные с этанолом, был в значительной степени неизвестен. В настоящем исследовании мы использовали мышь Glra2 KO, у которой отсутствует субъединица α2 глицинового рецептора, чтобы оценить вклад этой субъединицы в область, важную для обучения, основанного на вознаграждении.Используя эту мышиную модель, ранее был сделан вывод, что ген критически важен для развития мозга, а его делеция вызвала дефицит памяти распознавания объектов и нарушение LTP в префронтальной коре (Pilorge et al., 2016). Изменение глутаматергической передачи, корковой синаптической пластичности (Morelli et al., 2017) и нарушение консолидации моторной памяти также были обнаружены у этой мыши (Molchanova et al., 2018), что свидетельствует о наличии компенсаторных механизмов. С другой стороны, с использованием другой модели мышей Glra2 KO было показано, что эта мышь демонстрирует нарушенный нейрогенез гиппокампа взрослых и дефицит пространственной памяти (Lin et al., 2017).
4.1 Роль глициновых рецепторов, содержащих субъединицу α2, в нейронах прилежащего ядра
Предыдущие исследования продемонстрировали экспрессию субъединиц α2 глицинового рецептора в прилежащем ядре (Jonsson et al., 2012; Jonsson, Kerekes, Hyytia, Ericson, & Soderpalm, 2009), и наши результаты согласуются с этими выводами. Например, вестерн-блоттинг мышей Glra2 — / Y показал более низкие уровни белка α-субъединиц глицинового рецептора в прилежащем ядре, в то время как анализ qPCR показал, что субъединица α2 полностью исчезла.Интересно, что результаты, полученные с помощью кПЦР, показали более высокую относительную экспрессию субъединиц α1 и α3 глицинового рецептора и более низкую относительную экспрессию субъединиц β глицинового рецептора у мышей Glra2 — / Y . Тем не менее, электрофизиологические записи в прилежащих нейронах показали, что функциональные рецепторы в основном отсутствовали в большинстве нейронов мышей Glra2 — / Y , что хорошо коррелирует с результатами вестерн-блоттинга. Кривые «концентрация глицина – ответ» для тех нейронов, которые ответили на глицин, показали значение EC 50 , которое предполагает присутствие субъединиц α1 рецептора глицина, поскольку это значение сопоставимо с EC 50 41 ± 1 мкМ для субъединицы рецептора глицина α1 полученные в клетках HEK293 (Yevenes et al., 2010). Возможное объяснение такого небольшого тока в присутствии даже высоких уровней мРНК α-субъединицы другого глицинового рецептора и белков β-субъединицы может быть основано на стабильности полученной мРНК или потенциальных доминантно-негативных эффектах мышей Glra2 KO. Подобный эффект наблюдался у мышей KO для субъединицы α6 рецептора GABA A , где удаление субъединицы α6 изменяло уровни субъединицы δ в гранулярных клетках мозжечка (Jones et al., 1997). Наши результаты, тем не менее, согласуются с предыдущими выводами, которые показали отсутствие активируемых глицином токов в полосатом теле у мышей Glra2 KO (McCracken et al., 2017; Молчанова и др., 2018), область, где α2 является преобладающей субъединицей у мышей WT (Comhair et al., 2018). Дефицит, однако, касался не только глицинергической функции, поскольку мы обнаружили, что амплитуда вызванных ГАМК токов была также снижена у мышей Glra2 — / Y .
Предыдущие отчеты показали наличие глицинергической нейротрансмиссии в мезолимбической системе, нечувствительной к этанолу (Muñoz et al., 2018).В настоящем исследовании мы обнаружили уменьшение количества нейронов с глицинергическими синаптическими токами у мышей Glra2 — / Y по сравнению с мышами WT. Интересно, что нейроны, которые отображали глицинергические mIPSC, также демонстрировали тенденцию к снижению частоты общих mPSC, что указывает на роль субъединиц рецептора глицина α2 в установлении ГАМКергических и глутаматергических входов в прилежащем ядре. Кроме того, мы обнаружили небольшое увеличение значения постоянной времени распада в нейронах, обладающих глицинергическими IPSC, у мышей Glra2 — / Y , что указывает на изменения в конформации синаптических рецепторов глицина, где могут находиться как α1-, так и α3-субъединицы. преобладающие формы, согласно анализу КПЦР.Несмотря на сниженную глицинергическую синаптическую нейротрансмиссию в прилежащем ядре мышей Glra2 — / Y , не было изменений в уровнях субъединицы β глицинового рецептора, как показал вестерн-блот-анализ. Поскольку субъединица β необходима для кластеризации рецепторов глицина в постсинаптических сайтах (Meyer, Kirsch, Betz, & Langosch, 1995) и не образует функциональных гомомерных каналов (Grudzinska et al., 2005), эта субъединица в Glra2 — / Y мышь, вероятно, остается в нефункциональном состоянии, что объясняет сокращение ввода.
Взятые вместе, эти результаты демонстрируют важный вклад субъединиц рецептора глицина α2 в глицинергический компонент в прилежащих нейронах, поскольку глицинергические токи у мышей Glra2 — / Y были заметно снижены. С другой стороны, эти данные также предполагают, что субъединицы α2-глицинового рецептора могут быть важны для правильной экспрессии и функции α1- и α3-субъединиц рецептора глицина и рецепторов GABA A в прилежащем ядре.
4.2 Поведенческие эффекты высоких доз этанола у мышей
Glra2 KOПрилежащее ядро является важной входной структурой базальных ганглиев, которая интегрирует информацию от корковых и лимбических структур для обеспечения целенаправленного поведения (Scofield et al., 2016). Таким образом, хотя в нескольких исследованиях сообщалось, что у детенышей мышей, лишенных α2-субъединицы глицинового рецептора, постнатально наблюдалась микроцефалия (Avila et al., 2014), и что α2-глициновые субъединицы рецептора способствовали миграции кортикальных интернейронов (Avila et al., 2013), взрослые мыши Glra2 — / Y , использованные в этом исследовании, не имели различий в массе мозга или общей массе и не демонстрировали увеличения мышечного тонуса. Это согласуется с предыдущими отчетами, которые показали, что этот пониженный вес мозга постепенно ослабевает с возрастом (Morelli et al., 2017) и устраняется в зрелом возрасте (Pilorge et al., 2016). Несмотря на то, что эта животная модель является достаточно нормальной и может позволить нам узнать о механизмах, связанных с потреблением алкоголя, широкое влияние на развитие мозга и связанные с ним компенсаторные адаптации множества других рецепторов нейротрансмиттеров в некоторой степени препятствуют приписыванию более конкретных поведенческих результатов нормальной роли. субъединицы α2, или действия алкоголя конкретно на субъединицу α2.
Анализв открытом поле не выявил различий между генотипами в общем пройденном расстоянии или времени, проведенном в центре, что согласуется с ранее опубликованными данными (Blednov et al., 2015; Lin et al., 2017; Molchanova et al., 2018; Pilorge et al., 2016). Однако мы обнаружили изменения в навыках моторного обучения с использованием ускоряющего вращающегося стержня, который показал увеличенную задержку падения как у самцов, так и у самок мышей Glra2 KO. Хотя этот результат не был обнаружен в предыдущих исследованиях (Молчанова и др., 2018; Pilorge et al., 2016), это может быть связано с различиями в протоколе, поскольку мы тестировали мышей в течение двух дней подряд с пятью испытаниями в день 1 и 10 испытаниями на день 2, в то время как Молчанова и соавт. тестировали мышей в течение пяти дней подряд с четырьмя испытаниями на каждый день. Повышенная латентность падения, обнаруженная у мышей Glra2 KO, может быть объяснена компенсацией нейронов в базальных ганглиях, важной области для обучения двигательным навыкам.
Острое употребление опьяняющих доз этанола вызывает быстрые изменения функций мозга от нарушения координации, моторной атаксии и седативного эффекта до угнетения дыхания, комы и смерти при приеме более высоких доз этанола.С другой стороны, его хроническое употребление связано с стремлением к алкоголю, пьянством, толерантностью и зависимостью (Spanagel, 2009). Чтобы начать понимание роли субъединиц рецептора глицина α2 в поведении, связанном с опьяняющими дозами этанола, мы оценили атаксию и седативный эффект, вызванные этанолом. Хотя время восстановления после атаксии с использованием анализа с фиксированным вращающимся стержнем не отличалось между генотипами как у мужчин, так и у женщин, мы обнаружили различия в начале и продолжительности потери рефлекса выпрямления у мужчин.Кроме того, сходная концентрация этанола в крови, обнаруженная у обоих генотипов, показала, что это различие в седативном действии не было связано с фармакокинетикой этанола. Уменьшение продолжительности потери рефлекса выпрямления у самцов мышей Glra2 — / Y выявило важность субъединицы α2 в регуляции пробуждения, седативного эффекта и потери сознания. Важно отметить, что уменьшенная продолжительность потери рефлекса выпрямления, обнаруженная у мышей Glra2 — / Y , наблюдалась в двух независимых лабораториях, что подтверждает результаты, полученные с использованием этой линии KO.Кроме того, эти результаты согласуются с неопубликованными результатами модели KI на мышах для субъединицы α2 рецептора глицина, в которой остатки лизина и аргинина были заменены на аланины в положениях аминокислот в большой внутриклеточной петле, соответствующей KK385–386 в глицине. субъединица рецептора α1 (рукопись готовится). Кроме того, уменьшение продолжительности потери рефлекса выпрямления согласуется с идеей о том, что этанол потенцирует функцию рецептора глицина, что приводит к эффекту седативного эффекта (Burgos et al., 2015). Удивительно, но мы не обнаружили различий в начале или продолжительности утраты рефлекса выпрямления между генотипами у самок мышей. Тем не менее, важно отметить, что самки мышей WT показали меньшую продолжительность потери рефлекса выпрямления по сравнению с самцами мышей WT, как и в предыдущих сообщениях (Blednov et al., 2003). Мы объяснили это различие механизмами, связанными с полом у женщин. Например, в предыдущих исследованиях сообщалось о половых различиях во времени сна, вызванных этанолом (DeFries et al., 1989) и подразумевает действие нейростероидов (King, 2008; Morrow, VanDoren, Fleming, & Penland, 2001). Эти данные подчеркивают, что, хотя субъединица рецептора глицина α2 вносит важный вклад в некоторые эффекты этанола, такие как потеря рефлекса выпрямления, это поведение является более сложным и, вероятно, затрагивает несколько мишеней.
4.3 Изменения в потреблении и предпочтении этанола
Расстройство, связанное с употреблением алкоголя, — это рецидивирующее заболевание головного мозга с серьезными социальными последствиями, точный механизм которого до сих пор неизвестен.Управлением по контролю за продуктами и лекарствами США для лечения расстройств, связанных с употреблением алкоголя, одобрено всего три препарата (дисульфирам, налтрексон и акампросат) (Mann, 2004; Spanagel, 2009). Пьянство — это форма злоупотребления алкоголем, определенная Национальным институтом злоупотребления алкоголем и алкоголизмом как употребление алкоголя для достижения концентрации этанола в крови не менее 80 мг · дл -1 . Обычно это происходит примерно через 2 часа после четырех порций для женщин и пяти порций для мужчин. Хотя пьянство — это не то же самое, что и расстройство, связанное с употреблением алкоголя, оно значительно увеличивает риск развития проблемы злоупотребления алкоголем (Gowin, Sloan, Stangl, Vatsalya, & Ramchandani, 2017).Повторяющиеся циклы запоя и воздержания являются ключевыми компонентами в развитии зависимости. Мы использовали парадигму питья в темноте, чтобы оценить, есть ли разница в структуре потребления между мышами WT и мышами Glra2 KO. Важной особенностью этой процедуры является то, что мыши самостоятельно вводят алкоголь и достигают концентраций этанола в крови до уровней алкогольного поведения (Rhodes et al., 2005; Rhodes et al., 2007). Испытание на питье в темноте показало более высокое потребление в дни 1 и 2 у самцов мышей Glra2 — / Y по сравнению с мышами WT, что выявило уровни концентрации этанола в крови выше запоя.Интересно, что не было различий в потреблении на 4-й день между обоими генотипами. Мы полагаем, что этот потолочный эффект на потребление этанола представляет собой тип гомеостатического регулирования потребления, при котором предыдущая интоксикация ограничивает последующее потребление. Для мышей KO они достигают уровня запоя во время первого воздействия, тогда как WT достигают уровня запоя на четвертый день (рис. 6b). Предыдущие исследования показали аналогичную картину потребления у мышей KI субъединицы α1 глицинового рецептора (Muñoz et al., 2019), а недавние неопубликованные результаты демонстрируют такое же питьевое поведение на мышиной модели KI субъединицы α2 глицинового рецептора (рукопись в стадии подготовки), подтверждая вывод о том, что рецепторы глицина играют решающую роль в потреблении этанола. Неожиданно мы обнаружили, что высокое потребление этанола у самок мышей WT не отличалось от мышей Glra2 — / — в день 1, но затем их потребление со временем стало одинаковым. Однако важно отметить, что самки мышей WT потребляли больше этанола, чем самцы мышей WT, поведение, которое можно объяснить тем фактом, что самкам требуется более высокая концентрация этанола в крови для достижения того же эффекта этанола, что отражается в потере тестов рефлекса выпрямления. .Этот результат подтверждает идею о достижении гомеостатическим контролем потолка потребления этанола, ведущего к интоксикации.
Предыдущие исследования сообщили о половых различиях в потреблении алкоголя с использованием грызунов в нескольких тестах на потребление этанола (Becker & Koob, 2016; Caruso et al., 2018; Rhodes et al., 2007) и продемонстрировали, что физиология яичников способствует увеличению потребления этанола за счет женщины (Satta, Hilderbrand, & Lasek, 2018). Кроме того, было продемонстрировано, что 17-β-эстрадиол ингибирует рецепторы глицина (Jiang et al., 2009), предполагая различную модуляцию глициновых рецепторов по полу, что может объяснить различия, обнаруженные в поведении самцов и самок мышей. С другой стороны, когда мы исследовали предпочтение условного места, мы обнаружили, что мыши Glra2 — / Y не подвергались воздействию этанола после восьми сеансов кондиционирования, возможно, потому, что у мышей KO этанол менее полезен для создания кондиционирования. по сравнению с мышами WT (Cunningham et al., 2006). Следовательно, им нужно потреблять больше этанола при питье в темноте, чтобы получить положительный эффект.Другое объяснение состоит в том, что, хотя самцы мышей Glra2 — / Y утратили предпочтение этанолу, они будут потреблять большое количество алкоголя, потому что они могут быть менее чувствительны к отвращающему эффекту этанола. Эти результаты предполагают, что мышам Glra2 KO потребуется больше сеансов кондиционирования, чтобы продемонстрировать предпочтение стороне этанола по сравнению с мышами WT. С другой стороны, возможно, что из-за того, что у мышей KO обнаружен дефицит памяти (Lin et al., 2017; Pilorge et al., 2016), они могут не вспомнить сторону с положительным подкреплением. Результаты, полученные при предпочтении условного места и питье в темноте, продемонстрировали важные различия в мотивации и предпочтении этанола у мышей Glra2 KO, которые можно объяснить отсутствием субъединиц α2 или компенсаторными эффектами, которые могут возникать у животных KO.
Настоящие результаты предполагают, что субъединица α2 рецептора глицина, как субъединица α1 рецептора глицина, играет важную роль в схеме вознаграждения мезолимбической дофаминергической системы.Что касается предыдущих исследований с использованием другой линии КО с субъединицей GlyR α2, в которой изучали влияние на поведение этанола (Blednov et al., 2015), эти мыши имели другой генетический фон с меньшим количеством обратных скрещиваний с мышами C57BL / 6J по сравнению с нашей линией мышей. Кроме того, мыши Glra2 KO, использованные в исследовании Бледнова, были созданы путем делеции экзонов 6 и 7 (Young-Pearse, Ivic, Kriegstein, & Cepko, 2006), что отличалось от нашего подхода к мышам KO. Эти различия в линиях мышей могут объяснить расхождения, обнаруженные между двумя исследованиями с точки зрения потребления этанола и седативного эффекта.Хотя наши результаты могут показаться противоречащими вышеупомянутому исследованию, важно подчеркнуть, что существуют убедительные доказательства того, что накопительные рецепторы глицина регулируют потребление алкоголя (Lido et al., 2011; Molander et al., 2005; Molander, Lido, Lof, Ericson , & Soderpalm, 2007; Muñoz et al., 2019). Основываясь на наших данных, мы предполагаем, что рецепторы глицина защищают от употребления алкоголя, поскольку рецепторы глицина в прилежащем ядре способны снижать потребление этанола. Вероятный механизм этой регуляции заключается в том, что в нейронной цепи, включающей прилежащее ядро - вентральную тегментальную область, активация акумбальных рецепторов глицина вызывает ингибирование нейронов со средними шипами, вызывая возбуждающий ответ в пути обратной связи к вентральной тегментальной области, подавляя дофаминергические нейроны. .Также важно отметить, что наши поведенческие исследования проводились на мышах в возрасте от 8 до 12 недель, представляющих молодых животных-подростков, стадии, на которой наблюдается высокая предрасположенность к зависимости, особенно у людей (Bava & Tapert, 2010 ; Карузо и др., 2018; Варлинская, Спир, 2015).
Таким образом, настоящее исследование с использованием мышей α2 KO представляет собой комплексную, физиологическую и поведенческую характеристику эффектов этанола на животной модели, которая, по-видимому, демонстрирует тонкие компенсаторные эффекты, хотя и не смертельные и не заметные с неврологической точки зрения.Чтобы сделать более решительный вывод о роли этой субъединицы, мы в настоящее время изучаем мышиную модель α2 KI, которая не демонстрирует заметных компенсаций, и анализируем нокдаун субъединицы в определенных областях мозга с помощью аденоассоциированного вируса (AAV). Наконец, это исследование предоставляет доказательства того, что мыши с делецией в Glra2 являются полезной моделью не только для изучения расстройств аутистического спектра (Pilorge et al., 2016), но и для расстройств, связанных с употреблением алкоголя.
БЛАГОДАРНОСТИ
Мы благодарим Dr.Греггу Э. Homanics за комментарии к статье. Также мы благодарим Лорен Агуайо и Каролину Бенитес за техническую помощь.
Эта работа была поддержана грантом R01AA025718 Национального института здравоохранения (NIH), Национальным комитетом исследований и исследований (CONICYT), грантом DPI 20140008, грантом G0500833 Совета по медицинским исследованиям (MRC) и Фламандским исследовательским фондом (Fonds Fonds Fonds Wetenschap ]) 1518419N и 1519516N Федеральное управление научной политики Бельгии (BELSPO) Межуниверситетские полюса притяжения P7 / 10.
ВКЛАД АВТОРА
Л.С.М., С.Г., и Л.Г.А. участвовал в разработке дизайна. L.S.M., S.G., A.A., N.R., J.C., and G.M. провели эксперименты и проанализировали данные. R.J.H. построила линейку мышей Glra2 KO. L.S.M., L.G.A., R.J.H., J.M.R. и B.B. написали или участвовали в написании рукописи. Все авторы рецензировали рукопись.
КОНФЛИКТ ИНТЕРЕСОВ
Авторы заявляют об отсутствии конфликта интересов.
ДЕКЛАРАЦИЯ ПРОЗРАЧНОСТИ И НАУЧНОЙ СТЕПЕННОСТИ
В данной Декларации подтверждается, что в данной статье соблюдаются принципы прозрачной отчетности и научной строгости доклинических исследований, изложенные в руководстве BJP по дизайну и анализу, иммуноблоттингу, иммунохимии и экспериментам на животных, а также рекомендациям финансирующих агентств, издателей и другие организации, занимающиеся вспомогательными исследованиями.
новых данных о пользе для здоровья — ScienceDaily
На протяжении веков люди, которые чувствовали себя больными или находились в состоянии стресса, пробовали пить ромашковый чай как лекарство от всех болезней.Теперь исследователи в Англии нашли новые доказательства того, что популярный травяной чай может действительно помочь облегчить широкий спектр заболеваний, включая простуду и менструальные спазмы.
Их исследование планируется опубликовать 26 января в журнале Американского химического общества по сельскохозяйственной и пищевой химии, одном из рецензируемых журналов Общества. ACS — крупнейшее научное сообщество в мире.
«Это одно из растущего числа исследований, которые предоставляют доказательства того, что обычно используемые натуральные продукты действительно содержат химические вещества, которые могут иметь лечебное значение», — говорит руководитель исследования Элейн Холмс, доктор философии.D., химик из Имперского колледжа Лондона. «Отрасль здравоохранения уделяет все большее внимание функциональным продуктам питания, включая натуральные лечебные средства, однако мало исследований проводилось в отношении долгосрочного воздействия таких продуктов на биологию человека».
Травяным растением, используемым в этом исследовании, была немецкая ромашка (Matricaria recutita), также известная как мансанилла, цветы и листья которой заваривают как ароматный ароматный чай. В исследовании приняли участие четырнадцать добровольцев (семь женщин и семь мужчин), каждый из которых пил по пять чашек травяного чая ежедневно в течение двух недель подряд.Ежедневные пробы мочи брались и тестировались на протяжении всего исследования, как до, так и после употребления ромашкового чая.
Исследователи обнаружили, что употребление чая было связано со значительным повышением уровня гиппурата в моче, продукта распада некоторых растительных соединений, известных как фенолы, некоторые из которых связаны с повышенной антибактериальной активностью. По словам исследователей, это может помочь объяснить, почему чай укрепляет иммунную систему и борется с инфекциями, связанными с простудой.
Употребление чая также было связано с увеличением уровня глицина в моче — аминокислоты, которая, как было показано, снимает мышечные спазмы. Это может объяснить, почему чай помогает облегчить менструальные спазмы у женщин, вероятно, за счет расслабления матки, говорят исследователи. Ученые отмечают, что глицин действует как нервный релаксант, что также может объяснить, почему чай действует как мягкое седативное средство. Они добавляют, что для этой цели в магазинах продаются добавки с глицином.
Уровни гиппурата и глицина оставались повышенными в течение двух недель после того, как участники исследования перестали пить чай, что указывает на то, что соединения могут оставаться активными в течение довольно долгого времени, по словам исследователей. Они подчеркивают, что необходимы дополнительные исследования, прежде чем можно будет установить более определенную связь между чаем и его предполагаемой пользой для здоровья. Финансирование этого исследования было предоставлено Oxford Natural Products, фармацевтической, нутрицевтической и технологической компанией, расположенной в Оксфорде, Англия.
Американское химическое общество — некоммерческая организация, учрежденная Конгрессом США, в которую входят более 159 000 химиков и инженеров-химиков. Он издает многочисленные научные журналы и базы данных, созывает крупные исследовательские конференции и предоставляет образовательные, научные программы и программы карьерного роста в области химии. Его главные офисы находятся в Вашингтоне, округ Колумбия, и Колумбусе, штат Огайо.
История Источник:
Материалы предоставлены Американским химическим обществом . Примечание. Содержимое можно редактировать по стилю и длине.
Добавка триметилглицина — польза для здоровья, дозировка, побочные эффекты
Резюме триметилглицина
Основная информация, польза для здоровья, побочные эффекты, использование и другие важные сведения
Триметилглицин (TMG) представляет собой молекулу, которая по своей структуре представляет собой аминокислоту глицин с тремя присоединенными к ней метильными группами. Он известен как молекула «бетаина» («бетаин» — это категория молекул), но поскольку это был первый обнаруженный диетический бетаин (из свеклы), и это самая популярная молекула, называемая бетаином, термин «триметилглицин» ‘и’ бетаин ‘используются как синонимы.
Основными механизмами бетаина являются либо его использование в качестве донора метила, когда он либо напрямую отдает метильную группу для восстановления гомоцистеина до L-метионина (рассматривается как кардиопротекторный), либо увеличивает уровень S-аденозилметионина (SAMe) в организме или активный молекулы фолиевой кислоты, и эти двое могут затем передать метильные группы другим частям тела. Благодаря этому добавление бетаина может косвенно поддерживать метилирование всего тела и напрямую поддерживать снижение уровня гомоцистеина (что достоверно наблюдается после приема добавок в умеренных и высоких дозах.
Другой важный механизм заключается в том, что бетаин представляет собой осмолит или молекулу, которая перемещается в клетку и выходит из нее, чтобы влиять на ее статус гидратации. Подобно креатину, повышенные внутриклеточные концентрации бетаина способствуют гидратации клеток и устойчивости к стрессовым факторам.
Бетаин (также известный как TMG) представляет собой «осмолит», молекулу, регулирующую водный баланс в клетках. Бетаин может непосредственно метилировать гомоцистеин, обладающий кардиозащитным действием. Он также косвенно влияет на метаболизм фолиевой кислоты и SAMe, поддерживая метилирование всего тела.
Если посмотреть на данные о людях на данный момент, то окажется, что бетаин эффективен и надежен для снижения концентрации гомоцистеина при ежедневном приеме в дозе 3 г или более. Однократная доза бетаина снижает уровень гомоцистеина, который остается подавленным до тех пор, пока продолжается прием добавок. Было обнаружено, что бетаин снижает уровень гомоцистеина на 10% у людей с нормальным уровнем или на 20-40% у людей с повышенным уровнем гомоцистеина.
Известно, что уровень гомоцистеина повышен у лиц с проблемами сердечно-сосудистой системы, и он является биомаркером сердечно-сосудистых осложнений (т.е.когда уровень гомоцистеина повышен, субъект также подвергается более высокому риску возникновения проблем). Однако корреляция не указывает на причинно-следственную связь, поэтому неясно, является ли гомоцистеин просто индикатором проблем (биомаркером) или может действительно способствовать сердечнососудистой дисфункции. Считается, что снижение уровня гомоцистеина защищает сердце, но прямая связь между добавлением бетаина и защитой сердечно-сосудистой системы у людей еще не установлена.
В другом месте, хотя доказательства в настоящее время немного сбивают с толку (предварительные доказательства демонстрируют удивительные перспективы и одно контролируемое исследование не может воспроизвести его, но необходимы дополнительные исследования для подтверждения) бетаин может играть роль в лечении ожирения печени и связанной с ней печени. фиброз при приеме в высоких дозах (20 г в день).
Что касается здоровья, то бетаин наиболее перспективен для защиты печени и сердечно-сосудистой системы. Теоретически он должен обладать высокой защитой, но исследований, непосредственно оценивающих связь между добавлением бетаина и улучшенными биомаркерами здоровья, еще не существует.
Наконец, бетаин рекомендован в качестве соединения, повышающего эффективность, хотя и с весьма ненадежными результатами. При приеме 1,25 г два раза в день бетаин иногда был связан с увеличением выходной мощности (только с ошибкой в других случаях) и незначительным увеличением объема тренировки и выносливости (немного более надежным, чем выходная мощность, но все же не единодушное мнение).Общий размер эффекта или то, насколько бетаин приносит пользу субъекту, кажется довольно небольшим, и на данный момент все исследования, показывающие пользу от добавок бетаина, были связаны с Danisco (DuPont Nutrition), производителем бетаина.
Польза TMG для физических упражнений, даже если предполагается, что она присутствует, может иметь мало практического значения или не иметь никакого практического значения, поскольку:
-
Уровень бетаина в сыворотке повышен в той же степени с 1 г бетаина, что и с 1 г добавок холина, и последний может иметь некоторые центрально действующие (связанные с мозгом) преимущества при тренировках, при этом, возможно, дешевле
-
Теория, которая больше всего соответствует наблюдаемым преимуществам для физической работоспособности (действия осмолита, защищающие клетку), представляет собой механизм, который не только может повторяться с холином, как упоминалось ранее, но и механизм действия креатина; единственное исследование по оценке комбинации не выявило аддитивного эффекта бетаина на пользу креатина
Вы должны войти в систему, прежде чем сможете следить за этой страницей.
Вы подписались на Триметилглицин.